
1.4: Recorrido por la célula eucariota
- Page ID
- 54025

A. El Núcleo
El núcleo separa el plano genético, es decir, el ADN del citoplasma celular. Aunque el núcleo eucariota se descompone durante la mitosis y la meiosis a medida que se forman los cromosomas y las células se dividen, pasa la mayor parte de su tiempo en interfase, el tiempo entre divisiones celulares. Aquí es donde se regula el estado de los genes (y por lo tanto de las proteínas producidas en la célula). ARNr, ARNt y ARNm se transcriben a partir de genes, se procesan en el núcleo y se exportan al citoplasma a través de los poros nucleares. Algunos otros ARN permanecen en el núcleo, participando típicamente en la regulación de la actividad génica. En todos los organismos, las células en división deben producir y repartir copias de su material genético duplicado por igual entre nuevas células hijas. Veamos primero la organización estructural del núcleo, y luego su papel en la genética de la célula y de todo el organismo.
1. Estructura del Núcleo Interfásico
El núcleo es el orgánulo más grande de la célula. A continuación se muestra una imagen típica de microscopio electrónico de un núcleo, el orgánulo eucariota más grande de una célula.
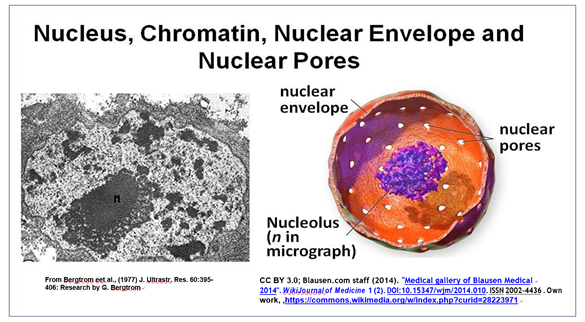
Esta sección transversal de un núcleo interfásico revela su doble membrana, o envoltura nuclear. La membrana externa de la envoltura nuclear es continua con el RER (retículo endoplásmico rugoso). Así, el lumen del RER es continuo con el espacio que separa las membranas de la envoltura nuclear. La micrografía electrónica también muestra un nucleolo prominente (marcado con n) y un RER oscuro granular que rodea el núcleo. Amplíe la micrografía; es posible que vea la doble membrana de la envoltura nuclear. También se pueden distinguir ribosomas (pequeños gránulos) unidos tanto al RER como a la membrana nuclear externa. Los poros de la envoltura nuclear (ilustrados en la caricatura de la derecha) permiten que moléculas grandes e incluso partículas se muevan dentro y fuera del núcleo a través de ambas membranas.
El núcleo no es un espacio desorganizado rodeado por la envoltura nuclear, como parece aparecer en las micrografías electrónicas de transmisión. El nucleolo es solo la mayor de varias inclusiones nucleares que parecen segregar funciones nucleares.
Santiago Ram ó n y Cajal reportó más estructuras en los núcleos de las neuronas hace más de 100 años, dibujando sus observaciones antes de que la tecnología fotomicrográfica moderna se pusiera ampliamente disponible. Vea lo que vio en los Cuerpos Nucleares de Cajal, incluyendo el nucleolo y lo que llegó a conocerse como cuerpos Cajal (CB). Como vimos antes, Ramón y Cajal compartieron el Premio Nobel de Fisiología o Medicina 1906 con Camillo Golgi por sus estudios de estructura de células nerviosas. Echa un vistazo a una galería de micrografías dibujadas a mano de Cajal de células nerviosas cerebrales en Beautiful Brain Cells de Cajal.
Más tarde vistos en un microscopio electrónico, los CB parecen bobinas de hilo enredado, y así fueron llamados cuerpos enrollados (convenientemente, también CB). Otros cuerpos nucleares desde que se identificaron incluyen Gemas, cuerpos PML, motas nucleares (o motas de empalme), cuerpos de locus de histonas (HLB)..., ¡y más! Diferentes cuerpos nucleares resultan estar asociados con proteínas específicas. La localización de proteínas específicas a diferentes cuerpos nucleares se puede observar en la micrografía de inmunofluorescencia a continuación.
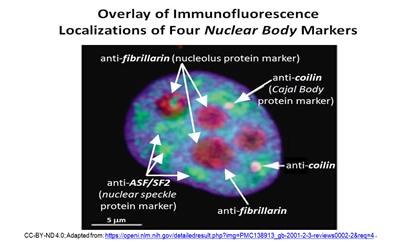
Los nucleolos contienen proteínas fibrilarinas y se tiñen de rojo porque han sido tratados con anticuerpos antifibrilarina marcados con fluorescencia roja. Los CBs contienen la proteína coilina. Fluorescen rosa porque los núcleos fueron tratados con anticuerpos anticoilina etiquetados con fluorescencia. Los anticuerpos fluorescentes verdes contra la proteína ASF/SF2 se localizan en motas nucleares. Como parte de, o incluidos en una matriz nuclear, los cuerpos nucleares organizan y regulan diferentes aspectos de la actividad nuclear y la función molecular. Los diferentes cuerpos nucleares realizan funciones específicas e interactúan entre sí y con las proteínas ADN y ARN para hacerlo. Volveremos a visitar algunos cuerpos nucleares en su contexto de trabajo en capítulos posteriores.
2. Cada célula (es decir, cada núcleo) de un organismo contiene los mismos genes
Leemos anteriormente que las bacterias están ocupadas duplicando y dividiendo sus cromosomas de ADN desnudos al mismo tiempo que crecen y se dividen por fisión binaria. En las células eucariotas, un ciclo celular divide la vida en eventos consecutivos discretos. Durante la mayor parte del ciclo celular, las células están en interfase y el ADN está envuelto en proteínas en una estructura llamada cromatina. No es simplemente el ADN, sino la cromatina la que debe duplicarse cuando las células se reproducen. La duplicación de ADN implica reordenar las proteínas de la cromatina. Esto ocurre antes de la división celular (mitosis y citocinesis). A medida que se acerca el tiempo de división celular, la cromatina se asocia con aún más proteínas, condensándose para formar cromosomas, mientras que la envoltura nuclear se disuelve.
Tal vez recuerdes que cada célula somática de un organismo contiene cromosomas homólogos emparejados, y por lo tanto dos copias de cada gen que posee un organismo. Por otro lado, los espermatozoides y óvulos contienen uno de cada par de cromosomas, y así una copia de cada gen. Ya sea por mitosis o meiosis, la citocinesis separa los cromosomas duplicados en células hijas. En la micrografía de fluorescencia de una célula en la etapa metafásica de la mitosis (abajo), los cromosomas (azul) están a punto de ser separados por los microtúbulos del aparato huso (verde).
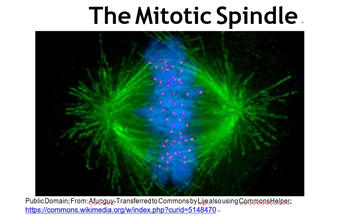
A medida que los cromosomas se separan y se forman las células hijas, los núcleos reaparecen y los cromosomas se descondensan. Estos eventos marcan la principal diferencia visible entre la división celular en bacterias y eucariotas. La citocinesis comienza cerca del final de la mitosis. La reproducción sexual, una característica clave de los eucariotas, implica meiosis más que mitosis. El mecanismo de la meiosis, la división de las células germinales que conduce a la producción de espermatozoides y óvulos, es similar a la mitosis excepto que las últimas células hijas tienen solo uno de cada uno de los cromosomas parentales, eventualmente para convertirse en los gametos (óvulos o espermatozoides).
Un mensaje clave para llevar a casa aquí es que cada célula de un organismo multicelular, ya sea óvulo, esperma o somática, contiene el mismo genoma (genes) en su núcleo. Esto se entendió ya que la mitosis y la meiosis se describieron por primera vez a finales del siglo XIX. Sin embargo, finalmente se demostró en 1962, cuando John Gurdon y Shinya Yamanaka trasplantaron núcleos de las células intestinales a la rana Xenopus laevis a huevos enucleados (huevos de los que se había extraído su propio núcleo). Estos 'huevos' crecieron y se desarrollaron hasta convertirse en renacuajos normales, demostrando que no se pierden genes durante el desarrollo, sino que solo se expresan diferencialmente.
Volveremos a visitar la clonación de animales más adelante en este libro. Pero por ahora, basta con saber que Molly la rana clonada fue seguida en 1996 por Dolly, la primera oveja clonada, y luego otros animales, todos clonados a partir de huevos enucleados trasplantados con núcleos celulares diferenciados. Haga clic en Clonar Cuarteterra para conocer la historia de 60 Minutos de la clonación de Cuarteterra, ¡una yegua polo campeona cuyos clones también son campeones! Para sus primeros experimentos de clonación animal, Gurdon y Yamanaka compartieron el Premio Nobel 2012 de Fisiología o medicina.
B. Ribosomas
En el otro extremo del espectro de tamaño, los ribosomas son máquinas sintetizadoras de proteínas conservadas evolutivamente en todas las células. Consisten en una subunidad grande y una pequeña, cada una compuesta por múltiples proteínas y una o más moléculas de ARN ribosómico (ARNr). Los ribosomas se unen a moléculas de ARN mensajero (ARNm), moviéndose a lo largo del ARNm a medida que traducen palabras de código de 3 bases (codones) para unir aminoácidos en polipéptidos. Múltiples ribosomas pueden moverse a lo largo del mismo ARNm, convirtiéndose en un polirribosoma, traduciendo simultáneamente el mismo polipéptido codificado por el ARNm. La aparición granular del citoplasma en las micrografías electrónicas se debe en gran parte a la distribución ubicua de subunidades ribosómicas y polisomas en las células.
La siguiente ilustración muestra una 'cadena' de ribosomas, el poliribosoma o polisoma para abreviar.
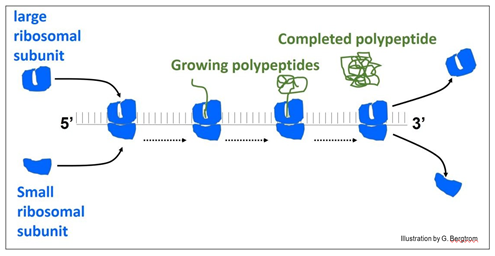
En la ilustración, los ribosomas se ensamblan a la izquierda del ARN mensajero (ARNm) para formar el polisoma. Cuando llegan al final del mensaje, los ribosomas se desensamblan del ARN y liberan el polipéptido terminado.
En una micrografía electrónica de células foliares de una planta de postre desecada quiescente, Selaginella lepidophylla, se pueden distinguir ribosomas distribuidos aleatoriamente y subunidades ribosómicas (flechas, abajo a la izquierda). En las células de una planta completamente hidratada, se pueden ver los polisomas como cadenas más organizadas de ribosomas (flechas, abajo a la derecha).

Los ribosomas eucariotas y procariotas difieren en el número de ARN y proteínas en sus subunidades grandes y pequeñas, y por lo tanto en su tamaño general. Los ribosomas aislados centrifugados en un gradiente de densidad de sacarosa se mueven a una velocidad basada en su tamaño (o más específicamente, su masa).
La siguiente ilustración muestra la diferencia en el 'tamaño' ribosómico, su composición proteica y el número y tamaños de sus ARN ribosómicos.
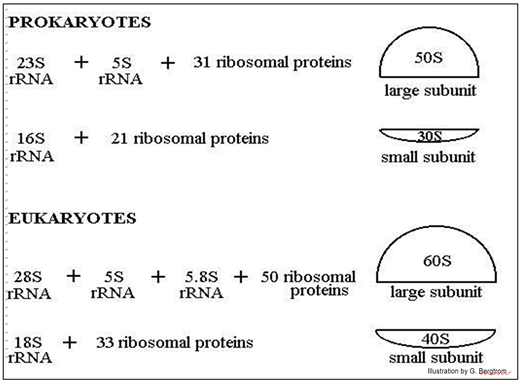
La posición de las subunidades ribosómicas en el gradiente está representada por un valor S, después de Svedborg, quien primero utilizó gradientes de densidad de sacarosa para separar macromoléculas y partículas por masa. Nótese que los propios ARN ribosómicos también se separan en gradientes de densidad de sacarosa por tamaño, de ahí sus diferentes valores de S.
C. Membranas internas y sistema endomembrano
Los microscopistas del siglo XIX vieron muchas de estas estructuras utilizando el arte de la histología, tiñendo células para aumentar el contraste visual entre las partes celulares. Uno de ellos, Camillo Golgi, un neurobiólogo temprano, desarrolló una mancha plateada (negra) que detectó por primera vez una red de vesículas que ahora llamamos cuerpos de Golgi (vesículas de Golgi) en las células nerviosas. Por sus descubrimientos en neurociencia celular, Golgi y Santiago Ramón y Cajal compartieron el premio Nobel de Medicina o Fisiología de 1906.
Muchas vesículas y vacuolas en las células, incluidas las vesículas de Golgi, forman parte del sistema endomembrano. Las proteínas sintetizadas en los ribosomas del RER (retículo endoplásmico rugoso) pueden ingresar al espacio interior (lumen) o pueden formar parte de la propia membrana del RER. La producción de RER, SER (retículo endoplásmico liso), cuerpos de Golgi, lisosomas, microcuerpos y otras membranas vesiculares, así como su contenido proteico, comienzan en el RER. El contenido de RER y proteína brota en vesículas de transporte que se fusionan con vesículas de Golgi (G en la micrografía electrónica a continuación).
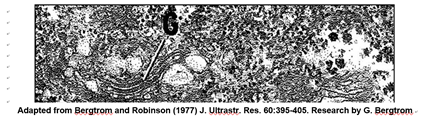
En su recorrido por el sistema endomembrano, las proteínas empaquetadas experimentan modificaciones escalonadas (maduración) antes de convertirse en biológicamente activas (abajo).
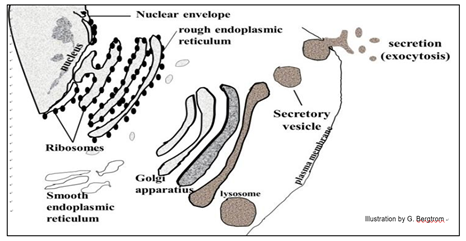
102 Vesículas de Golgi y el Sistema Endomembrana
Algunas proteínas elaboradas en el sistema endomembrano son secretadas por exocitosis. Otros terminan en orgánulos como los lisosomas que contienen enzimas hidrolíticas. Estas enzimas se activan cuando los lisosomas se fusionan con otros orgánulos destinados a la degradación. Las vacuolas de alimentos se forman cuando una membrana plasmática invagina, envolviendo partículas de alimentos. Luego se fusionan con lisosomas para digerir los nutrientes engullidos.
Los autofagosomas son pequeñas vesículas que rodean y eventualmente encapsulan orgánulos cansados (por ejemplo, mitocondrias desgastadas), fusionándose eventualmente con lisosomas cuyas enzimas degradan su contenido. En 2016, Yoshinori Ohsumi obtuvo el Premio Nobel de Fisiología y Medicina por casi 30 años de investigación desentrañando la biología celular y molecular de la autofagia. Los microcuerpos son una clase de vesículas más pequeñas que los lisosomas, pero formadas por un proceso similar. Entre ellos se encuentran los peroxisomas que descomponen los peróxidos tóxicos formados como subproducto de la bioquímica celular. Algunas vesículas que emergen del RER pasarán a formar parte del SER, que tiene varias funciones diferentes (por ejemplo, desintoxicación con alcohol en células hepáticas).
103 Retículo Endoplásmico Liso
Otros orgánulos incluyen las vacuolas contráctiles de protozoos de agua dulce que expulsan el exceso de agua que ingresa a las células por ósmosis. Algunos protozoos tienen extrusomas, vacuolas que liberan químicos o estructuras que disuaden a los depredadores o permiten la captura de presas. Una gran vacuola central acuosa domina el volumen de muchas células vegetales superiores. Cuando se llenan de agua, empujarán todas las demás estructuras contra la membrana plasmática. En una planta debidamente regada, esta vacuola llena de agua ejerce una presión osmótica que, entre otras cosas, evita que las hojas de las plantas se marchiten y los tallos erguidos.
D. Mitocondrias y Plastidios
Casi todas las células eucariotas contienen mitocondrias, que se muestran a continuación.
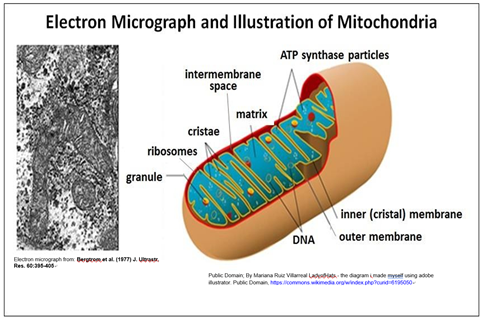
Una doble membrana rodea la mitocondria. Cada uno contiene y replica su propio ADN que contiene genes que codifican algunas proteínas mitocondriales. Obsérvese que el área superficial de la membrana mitocondrial interna se incrementa al ser plegada en cristae, que son sitios de respiración celular (oxidación aeróbica de nutrientes).
Anteriormente, especulamos con orgánulos eucariotas que podrían haberse originado dentro de las bacterias. Las mitocondrias probablemente evolucionaron a partir de una bacteria aeróbica completa (o proto-bacteria) que fue engullida por una célula eucariota primitiva. La bacteria escapó a la destrucción, convirtiéndose en endosimbionte en el citoplasma de la célula hospedadora. Lynn Margulis propuso por primera vez la Teoría Endosimbiótica (Margulis, L. [Sagan, L], 1967. Sobre el origen de las células mitosantes. Revista de Biología Teórica 14 (3): 225—274; disponible en: Margulis L. Teoría endosimbiótica). Margulis propuso que los cloroplastos también comenzaron como endosimbiontes. Al igual que las mitocondrias, los plastidios de las plantas y algunas algas tienen su propio ADN, muy probablemente originado como cianobacterias engullidas por células eucariotas primitivas. Viviendo en simbiosis con el resto de la célula, eventualmente evolucionarían a plastidios, incluidos los cloroplastos. La evidencia detallada de la Teoría Endosimbiótica se discute en otra parte.
Se encontró un puñado de protozoos carentes de mitocondrias y otros orgánulos. Esto había sugerido que podrían compartir ascendencia con aquellos eucariotas primitivos que adquirieron mitocondrias por endosimbiosis. Sin embargo, dado que tales células contienen otros orgánulos como hidrogenosomas y mitosomas, se piensa más probable que estas especies alguna vez tuvieron, pero luego perdieron mitocondrias. Por lo tanto, los descendientes de células eucariotas antiguas que carecen de mitocondrias probablemente ya no existan.
Los cloroplastos y cianobacterias contienen clorofila y utilizan un mecanismo fotosintético similar para producir glucosa. Un cloroplasto típico se muestra en la micrografía de abajo (izquierda). Un cloroplasto que comienza a almacenar azúcar nutriente como almidón está a la derecha.
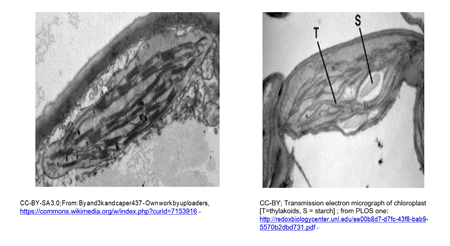
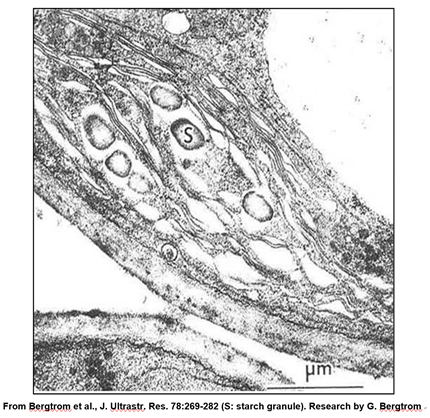
Un leucoplasto es un plastidio un cloroplasto que se ha llenado con gránulos de almidón. En la micrografía a continuación, se puede observar que, debido a la acumulación de almidón, los grana se han vuelto dispersos e indistintos, formando un leucoplasto.
E. Estructuras citoesqueléticas
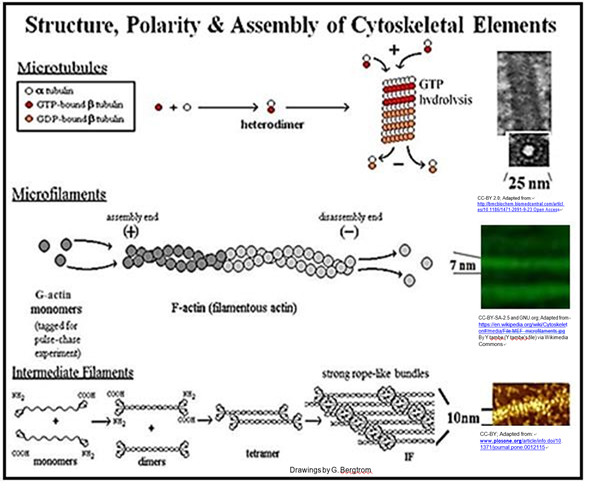
Hemos llegado a comprender que el citoplasma de una célula eucariota está altamente estructurado, permeado por bastones y túbulos. Los tres componentes principales de este citoesqueleto son microfilamentos, filamentos intermedios y microtúbulos.
Los microtúbulos están compuestos por monómeros de proteína a- y b -tubulina. Las proteínas monoméricas de actina conforman microfilamentos. Las proteínas de filamentos intermedios están relacionadas con la queratina, una proteína que se encuentra en el pelo, las uñas de las manos, las plumas de aves, etc. Las varillas y túbulos citoesqueléticos no solo determinan la forma celular, sino que también juegan un papel en la motilidad celular. Esto incluye el movimiento de las células de un lugar a otro y el movimiento de estructuras dentro de las células.
Ya hemos señalado que un citoesqueleto procariota está compuesto en parte de proteínas homólogas a actinas y tubulinas. Al igual que en un citoesqueleto eucariota, estas proteínas bacterianas pueden desempeñar un papel en el mantenimiento o cambio de la forma celular. Por otro lado, el movimiento impulsado por flagelos en bacterias se basa en la flagelina, una proteína que no se encuentra en las células eucariotas. Un flagelo bacteriano es en realidad una estructura rígida similar a un gancho unida a un motor molecular en la membrana celular que gira para propulsar la bacteria a través de un medio líquido.
En contraste, los microtúbulos eucariotas se deslizan uno junto al otro causando que un flagelo más flexible se ondula en movimientos ondulados. De igual manera, el movimiento de un cilio eucariota se basa en microtúbulos deslizantes, en este caso haciendo que los cilios latan en lugar de ondularse. Los cilios están involucrados no sólo en la motilidad, sino también en la alimentación y la sensación. A continuación se muestran las estructuras y el ensamblaje de los principales componentes citoesqueléticos.
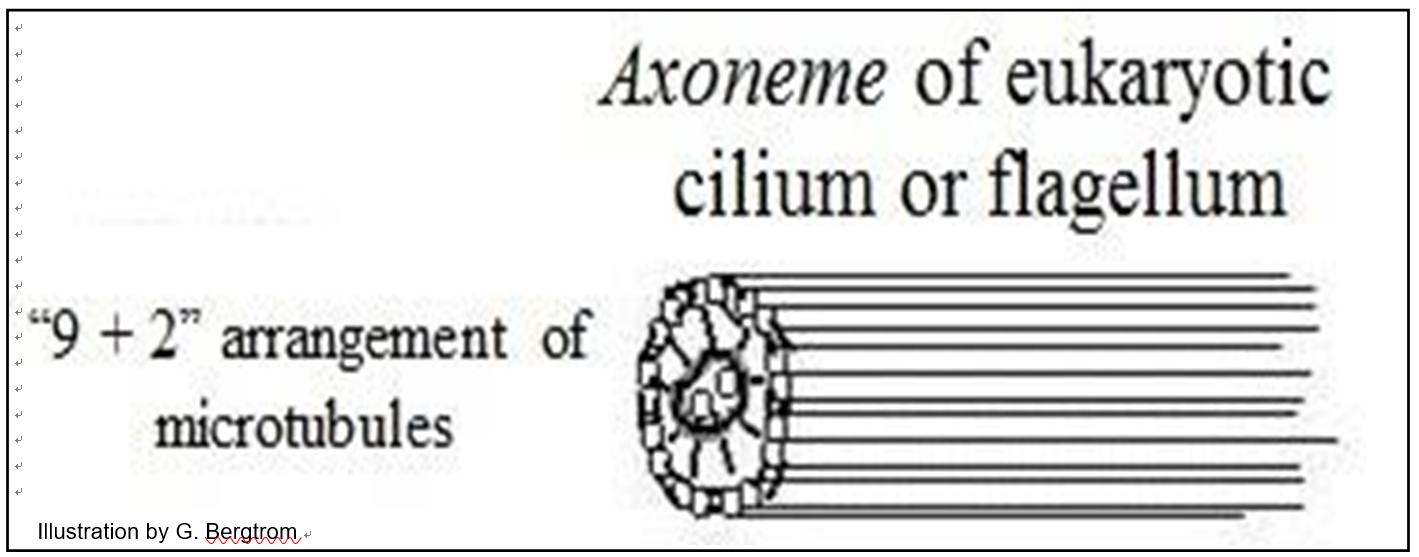
Los microtúbulos en flagelos eucariotas y cilios surgen de un cuerpo basal (similar a los cinetosomas o centriolos). Alineados en un flagelo o cilio, los microtúbulos forman un axonema rodeado por membrana plasmática. En micrografías electrónicas de secciones transversales, un axonema ciliar o flagelar se organiza típicamente como un anillo de nueve microtúbulos emparejados (llamados dobletes) alrededor de dos microtúbulos singlete (ilustrados a continuación).
Los centriolos están compuestos por un anillo de microtúbulos. En las células animales participan en la formación de fibras del huso durante la mitosis y son el punto desde el cual los microtúbulos irradian a través de la célula para ayudar a formar y mantener su forma. Estas estructuras no involucran axonemas. El aparato huso en células vegetales, que típicamente carecen de centriolos, se forma a partir de una estructura amorfa llamada MTOC, o Centro Organizador de Microtúbulos, que tiene el mismo propósito en mitosis y meiosis que los centriolos en las células animales.
106 Filamentos y Túbulos del Citoesqueleto
En otra parte, describimos cómo los microfilamentos y microtúbulos interactúan con las proteínas motoras (dineína, cinesina, miosina, etc.) para generar fuerza que resulta en el deslizamiento de filamentos y túbulos para permitir el movimiento celular. Verás que las proteínas motoras también pueden transportar moléculas de carga de un lugar a otro en una célula.