
18.3: La estructura molecular y la organización subcelular de los componentes citoesqueléticos
- Page ID
- 53872

De las tres principales fibras citoesqueléticas, los filamentos intermedios desempeñan un papel principalmente estructural en las células. Los microtúbulos y microfilamentos tienen funciones duales, manteniendo dinámicamente la forma celular y permitiendo la motilidad celular. Por ejemplo, cuando se unen a la membrana plasmática, los microfilamentos mantienen la forma celular. Sin embargo, al interactuar con proteínas motoras (por ejemplo, miosina), pueden tirar o empujar contra una membrana celular muscular, cambiando la forma de la célula. Asimismo, las proteínas motoras como la dineína y la cinesina pueden mover 'carga' de un lado a otro a lo largo de las huellas de los microtúbulos de un punto a otro en la célula. En breve veremos cómo interactúan las proteínas motoras con los microtúbulos y microfilamentos. En este punto, echemos otro vistazo a los dibujos y micrografías de los tres principales filamentos citoesqueléticos de células eucariotas (abajo) que vimos anteriormente en el texto.
321 Introducción al Citoesqueleto
La ubicación y las funciones generales de los microtúbulos, microfilamentos y filamentos intermedios se demostraron mediante microscopía de inmunofluorescencia. Después de exponer las células a anticuerpos marcados con fluoróforo contra proteínas de microtúbulos, microfilamentos (actina) o filamentos intermedios, las micrografías de fluorescencia de las células teñidas revelaron las diferentes ubicaciones de las fibras en las células. A continuación se muestra la localización típica de las diferentes fibras citoesqueléticas.
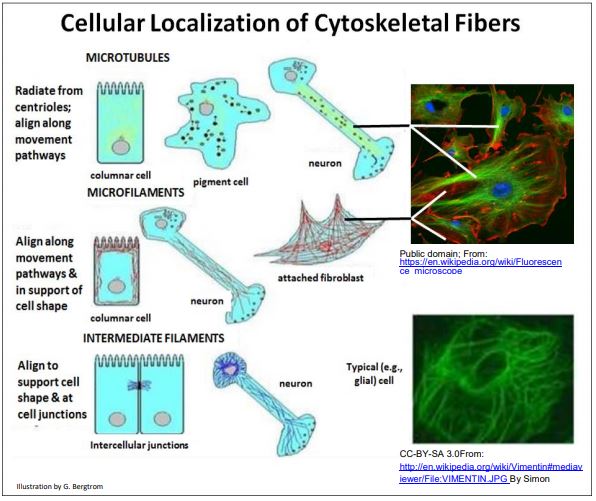
Estas localizaciones son consistentes con las funciones conocidas de los principales filamentos del componente citoesquelético en la estructura celular y la motilidad. A pesar del pequeño tamaño de las células procariotas, también recientemente se encontraron que tienen estructuras citoplásmicas previamente insospechadas que podrían servir como citoesqueleto (NCBI-a Procariota Cytoskeleton? ). Entonces, ¡quizás todas las células (no solo eucariotas) son más que una bolsa desorganizada de savia fluida! A continuación, consideramos roles específicos de microtúbulos, microfilamentos, filamentos intermedios y proteínas relacionadas en el citoesqueleto eucariota.
322 Microtúbulos, Microfilamentos y Filamentos Intermedios en Células
A. Microtúbulos - una visión general
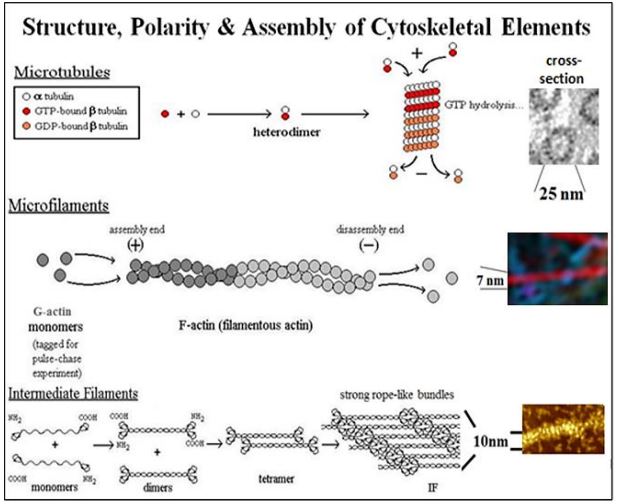
Los microtúbulos se ensamblan a partir de dímeros de monómeros de \(\alpha \)\(\beta \)-tubulina y -tubulina. Después de la formación, los dímeros \(\alpha \)/\(\beta \)-tubulina se agregan a un crecimiento, o extremo más (extremo +), alimentado por hidrólisis de GTP. El desmontaje en el extremo de los microtúbulos permite cambiar la forma de las células o la separación y movimiento de las cromátidas a polos opuestos de las células durante la división celular (es decir, mitosis o meiosis). Se demostró que los microtúbulos individuales aislados crecían por adición a un extremo y se desensamblaban en el extremo opuesto, distinguiendo así los extremos + y -extremos. Un resumen de este experimento que demuestra la polaridad de los microtúbulos se encuentra en el siguiente enlace.
323 Demostración de la Polaridad y Dinámica de Microtúbulos
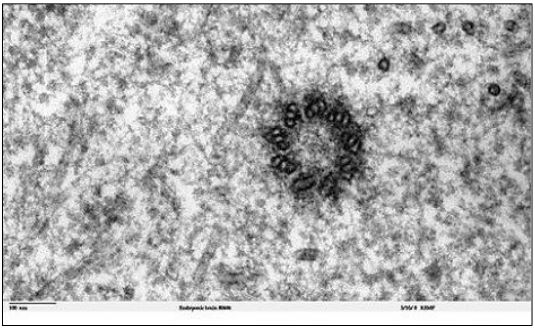
Los microtúbulos en la mayoría de las células pueden parecer desordenados. En interfase, tienden a irradiar desde centriolos en células animales que no se dividen, sin formar estructuras discretas. Sin embargo, en el período previo a la división celular, los microtúbulos se reorganizan para formar fibras de huso. Esta reorganización está nucleada a partir de centriolos en células animales y de un centro organizador de microtúbulos (MTOC) más amorfo en células vegetales. Un centríolo típico (o cuerpo basal) tiene una matriz de microtúbulos de '9 tripletes' como se ve en la sección transversal de micrografía electrónica (abajo).
1. Los dos tipos de microtúbulos en fibras de husillo
a) Microtúbulos de Cinetocoros
Los cromosomas duplicados se condensan en profase de mitosis y meiosis, formando cromátidas apareadas visibles unidas en sus centrómeros. Las proteínas específicas se asocian con los centrómeros para hacer un cinetocoro durante la condensación. A medida que se forma el aparato de huso, algunas fibras de huso se adhieren al cinetocoro; estos son los microtúbulos de cinetocoros. Por metafase, haces de microtúbulos de cinetocoros se extienden desde los cinetocoros en el centro celular hasta los centriolos polares o MTOC de la célula en división, como se dibuja a continuación.
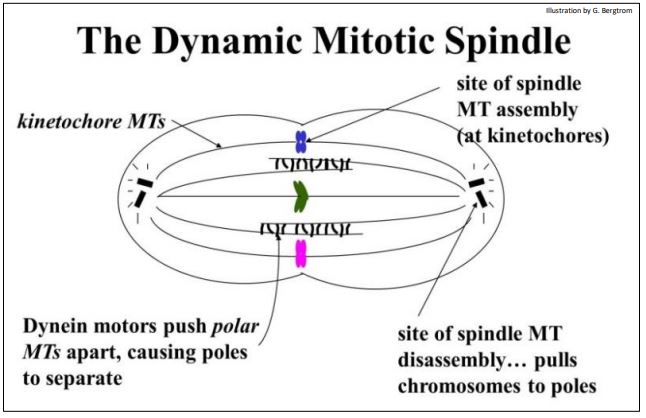
Ahora sabemos que los extremos +de los microtúbulos de cinetocoros están de hecho en los cinetocoros, ¡donde se ensamblan estas fibras! En la anafase, las fuerzas generadas cuando los microtúbulos se acortan en sus extremos —extremos (extremos de desmontaje) separan las cromátidas. El desmontaje de microtúbulos en los centrioles/MTOC proporciona la fuerza que atrae a los cromosomas hijos a los polos opuestos de la célula a medida que continúa la división celular.
b) Microtúbulos Polares
Los microtúbulos polares de fibra de huso se extienden desde centrioles/MTOC en polos opuestos de la celda. No se unen a cinetocoros de cromátidas, sino que se superponen en el centro de las células en división. A medida que los microtúbulos cinetocoros tiran de las cromátidas en la anafase, los microtúbulos polares se deslizan uno junto al otro en direcciones opuestas, separando los polos de la célula. En este caso, la dineína (una proteína motora unida a los microtúbulos) cataliza la hidrólisis de ATP para impulsar el deslizamiento de los microtúbulos. Los motores Dynein en los microtúbulos de un polo de la célula en efecto, 'caminan' a lo largo de microtúbulos superpuestos que se extienden desde el polo opuesto. El papel del desmontaje de microtúbulos en los centriolos (es decir, en el extremo negativo) se demostró en un ingenioso experimento en el que un pequeño rayo láser fue dirigido hacia una célula hacia fibras de huso unidas al cinetocoro de un par de cromátidas (ver esto animado en el siguiente enlace).
324 microtúbulos de fibra de husillo generan fuerza en cromátidas
2. Microtúbulos en Cilios y Flagelos
Los microtúbulos de cilios o flagelos emergen de un cuerpo basal, que se muestran a la izquierda en la micrografía electrónica de abajo
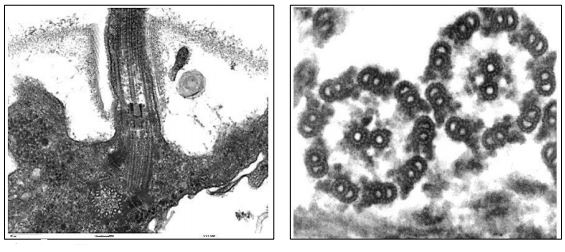
Los cuerpos basales son estructuralmente similares a los centriolos, organizados como un anillo de nueve trillizos de microtúbulos. La formación de cilios y flagelos comienza en los cuerpos basales pero muestran una disposición típica de 9+2 (9 dobletes externos más 2 microtúbulos centrales) en sección transversal (mostrada en la micrografía, arriba a la derecha). Después del tratamiento con detergente para eliminar las membranas de cilios o flagelos aislados, los axonemas restantes conservan la disposición de 9+2 microtúbulos. La relación estructural entre los axonemas de un cilio o flagelo y un microtúbulo individual se muestra en las secciones transversales siguientes.
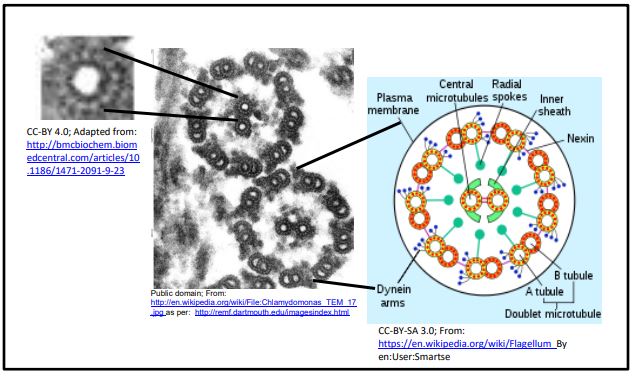
Es posible ver las subunidades de tubulina que componen un polímero de microtúbulos en sección transversal. Cada túbulo está formado por un anillo de 13 subunidades de tubulina. Los microtúbulos en los 'dobletes' comparten tubulinas, pero también están compuestos por 13 tubulinas. Cuando están completamente formados, los microtúbulos de 25 nm de diámetro parecen ser un cilindro hueco. Cuando se aíslan los microtúbulos, suelen venir junto con proteínas motoras de dineína y otras Proteínas Asociadas a Microtúbulos (MAP), algunas de las cuales mantienen los microtúbulos juntos en un axonema.
3. Las proteínas motoras de microtúbulos mueven la carga de un lugar a otro en las células
Las proteínas motoras como la dineína y la cinesina, son ATPasas; utilizan la energía libre de la hidrólisis de ATP para potenciar la motilidad intracelular. Echemos un vistazo más de cerca a cómo estas dos principales proteínas motoras transportan carga de un lugar a otro dentro de las células. Los orgánulos son una carga típica. Los ejemplos incluyen vesículas formadas en la cara trans del Golgi que contienen proteínas secretoras, pigmentos o neurotransmisores. Las vesículas secretoras se mueven a lo largo de las huellas de los microtúbulos hacia la membrana plasmática para exocitosis Las vesículas que contienen neurotransmisores se mueven desde el cuerpo celular de las neuronas a lo largo de las huellas de los microtúbulos en los axones, llegando a la terminación nerviosa donde se convierten en vesículas sinápticas. En un camaleón, las vesículas pigmentarias en las células de la piel se dispersan o agregan a lo largo de las huellas de los microtúbulos para cambiar el color de la piel
Las proteínas motoras transportan vesículas de carga en direcciones opuestas. El transporte de neurotransmisores es un ejemplo bien entendido. Las vesículas neurotransmisoras surgen del sistema endomembrano en cuerpos celulares neuronales. Impulsada por ATP, la cinesina potencia el movimiento de las vesículas anterógradas desde el cuerpo celular hasta las terminaciones nerviosas. En contraste, un motor de dineína dependiente de ATP, como parte de un complejo de dinactina, impulsa el movimiento retrógrado de las vesículas vacías de regreso al cuerpo celular. La estructura y acción de la proteína motora están por debajo.
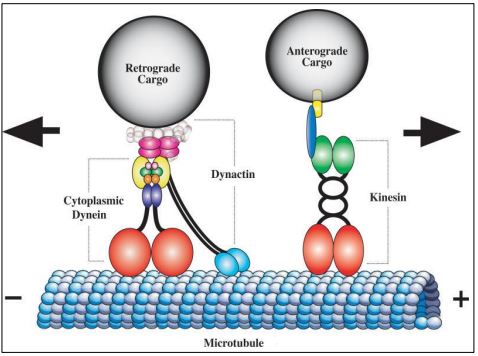
Un fantasioso (¡y no demasiado inexacto!) caricatura de una proteína motora 'caminando a lo largo de un microtúbulo axonal se anima en este enlace: Kinesin 'caminando' un orgánulo a lo largo de un microtúbulo.
325 Proteínas motoras de microtúbulos
En este punto, podemos observar varios tipos específicos de motilidad celular que involucran microtúbulos y microfilamentos.
4. La proteína motora Dynein permite que los axonemas se doblen
Echa un vistazo a la sección transversal de axonemas hace algunas ilustraciones. En el axonema 9+2 de cilios y flagelos, los brazos de dineína unidos a los túbulos A de los dobletes externos caminan a lo largo de los túbulos B del doblete adyacente. Si solo los dobletes de un lado de un axonema dan un paseo mientras los del otro lado se mantienen quietos, los microtúbulos se deslizarán uno junto al otro y el axonema (y por lo tanto un cilio o flagelo) se doblará. Sin embargo, el deslizamiento de los microtúbulos está limitado por nexin flexibles y accesorios de radios radiales. A continuación se ilustran los movimientos de cilios y flagelos.
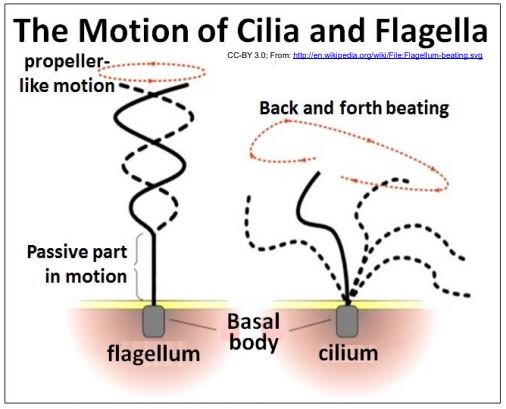
Las diferencias en el movimiento flagelar (hélice ondulada) y el movimiento ciliar (plano único, latido hacia adelante y hacia atrás) resultan en parte del cual los microtúbulos se deslizan en un momento dado y la naturaleza de su restricción por las proteínas axonémicas. Veamos algunos experimentos que demuestran estos eventos. CC-BY 3.0; De: es.wikipedia.org/wiki/Archivo:fl... um-beating.svg CMB3e 467.
Los experimentos en axonemas aislados demuestran el mecanismo de microtúbulos deslizantes de la motilidad ciliar y flagelar. En un experimento, se demostró que flagelos aislados y axonemas purificados “batieron” en presencia de ATP agregado (abajo).
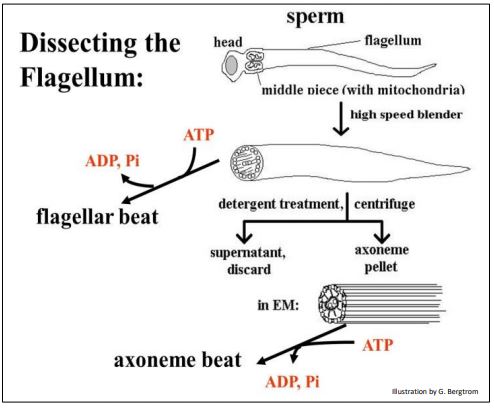
Agitar espermatozoides o células ciliadas en una licuadora de alta velocidad durante unos segundos cortará y separará flagelos o cilios del resto de la célula. Agregar ATP a los cilios o flagelos desprendidos hará que latan, un fenómeno que se ve fácilmente en un microscopio óptico. Los axonemas aislados de cilios o flagelos desprendidos mediante tratamiento con detergente (para romper las membranas) conservan su disposición característica de microtúbulos 9+2 así como otras características ultraestructurales..., ¡e incluso 'batirán' en presencia de ATP!
326 9+2 Matriz de microtúbulos en axonemas que superan
El tratamiento detergente adicional elimina los radios radiales, nexina y otras proteínas del axonema, provocando que los microtúbulos se separen. Luego se pueden observar dobletes de microtúbulos disociados y 'singletes' centrales en el microscopio electrónico.
Cuando se dializan los microtúbulos separados para eliminar los detergentes, los microtúbulos dobletes se vuelven a asociar, formando láminas, como se muestra en la caricatura a continuación.

El ATP agregado a estos dobletes de microtúbulos 'reconstituidos' hace que los microtúbulos se separen a medida que se hidroliza el ATP. Cuando tales preparaciones se fijan para microscopía electrónica inmediatamente después de agregar el ATP, quedan atrapadas en el acto de deslizamiento. Vea esto animado en el primer enlace a continuación.
327 Prueba de Microtúbulos Deslizantes Durante la Flexión de Flagelos y Cilios
328 flagelos bacterianos son alimentados por un gradiente de protones
329 Los efectos de diferentes fármacos sobre los microtúbulos y el cáncer
C. Microfilamentos - Estructura y Papel en la Contracción Muscular
A 7 nm de diámetro, los microfilamentos (filamentos de actina) son el componente citoesquelético más delgado. Los monómeros globulares de actina (G-actina) polimerizan para formar polímeros lineales de F-actina. Dos polímeros se combinan para formar el microfilamento de actina de doble espiral. Al igual que con los microtúbulos, los microfilamentos tienen un extremo +al que se agregan nuevos monómeros de actina para ensamblar F-actina, y un extremo en el que se desensamblan cuando están en un estado dinámico, como cuando una célula está cambiando de forma. Cuando un extremo de un microfilamento se ancla a una estructura celular, por ejemplo a placas en la membrana celular, las proteínas motoras como la miosina pueden usar ATP para generar una fuerza que deforme la membrana plasmática y así, la forma de la célula. Uno de los ejemplos mejor estudiados de interacción miosina/actina es en el músculo esquelético donde el deslizamiento de varillas de miosina gruesas altamente organizadas y los microfilamentos delgados de actina resulta en contracción muscular.
1. Filamentos Gruesos y Delgados de la Contracción del Músculo
Los haces de células musculares paralelas forman un músculo esquelético. Microscopía óptica de secciones delgadas de músculo esquelético muestran células musculares estriadas (miocitos, abajo).
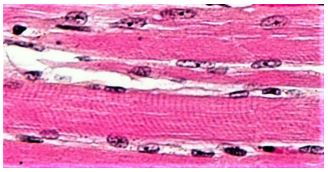
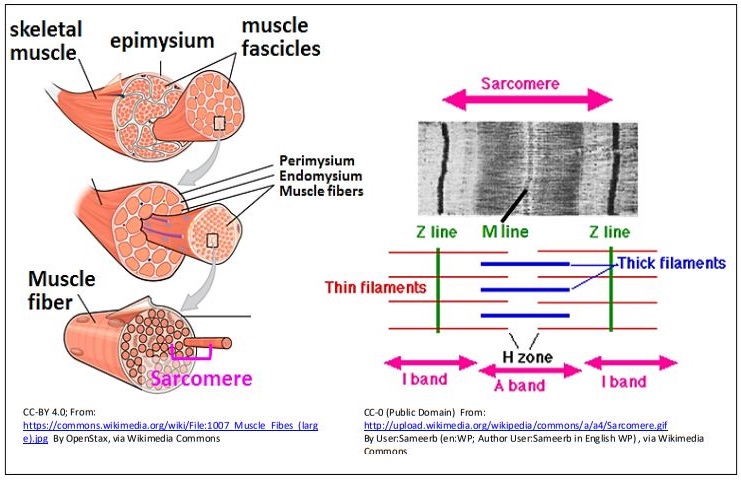
Las estructuras de color púrpura oscuro que rodean al miocito son las mitocondrias, que proporcionarán el ATP para alimentar la contracción. El músculo esquelético está formado por miocitos agrupados 'alineados'. Los miocitos agrupados (también llamados miofibras) se organizan aún más en fascículos que finalmente se agrupan en un músculo. La ilustración de reventón de la página siguiente muestra esta organización anatómica y estructura fina de un músculo (panel izquierdo).
La microscopía electrónica de alta resolución de la década de 1940 reveló la fina estructura del músculo esquelético (panel derecho de la ilustración), permitiendo la caracterización del sarcómero. Las bandas oscuras de las estrías en la micrografía ligera de los miocitos son regiones de sarcómeros alineados adyacentes. Un par de líneas Z demarcan un sarcómero (Z para zwischen, alemán para entre). La banda I es una región relativamente clara del sarcómero, compuesta principalmente por microfilamentos delgados (actina). La banda A en el centro del sarcómero consiste en superponer filamentos delgados y gruesos (actina y miosina), mientras que la zona H es una región donde la miosina no se solapa con filamentos de actina. Una línea M se encuentra en el centro de la zona H. Múltiples sarcómeros repetitivos de miocitos alineados en registro en los fascículos dan la apariencia de estrías en músculos enteros.
2. El modelo de filamento deslizante de la contracción del músculo esquelético
La microscopía electrónica del músculo relajado y contraído que se muestra a continuación es consistente con el deslizamiento de filamentos gruesos y delgados durante la contracción. Estructuras clave adicionales del sarcómero se pueden ver en el dibujo de la derecha.

Obsérvese que en los sarcómeros de una célula muscular contraída, la zona H casi ha desaparecido. Si bien el ancho de la banda A no ha cambiado después de la contracción, el ancho de las bandas I ha disminuido y las líneas Z están más cerca en el sarcómero contraído. La mejor explicación aquí fue la Hipótesis de Filamento Deslizante (modelo) de la contracción del músculo esquelético.
330 El modelo de filamento deslizante de contracción muscular esquelética
3. La paradoja de la contracción: la contracción y la relajación requieren ATP
El papel del ATP en alimentar el movimiento de los filamentos deslizantes durante la contracción del músculo esquelético se basó en parte en experimentos con fibras glicerinadas (fibras musculares empapadas en glicerina para permeabilizar la membrana plasmática). Los componentes citoplásmicos solubles se escapan de las fibras glicerinadas, pero dejan intactas las estructuras del sarcómero, tal como se visualiza por microscopía electrónica. Los investigadores encontraron que, si se volvía a agregar ATP y calcio a las fibras glicerinadas, el ATP se hidrolizaba y la fibra aún podría contraerse... ¡e incluso levantar un peso!
A continuación se ilustra la contracción de una fibra muscular glicerinada en presencia de ATP.
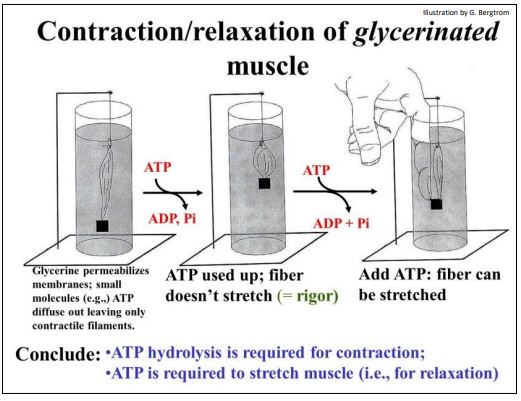
Cuando los ensayos mostraron que todo el ATP agregado había sido hidrolizado, el músculo permaneció contraído. ¡No se relajaría, incluso con el peso que había levantado todavía pegado! Intentar forzar manualmente el músculo hacia atrás a su posición relajada no funcionó. ¡Pero la fibra podría estirarse cuando se agregaba ATP fresco a la preparación! Además, si el experimentador soltaba inmediatamente después de estirar la fibra, ¡nuevamente se contraería y levantaría el peso! Se pudo repetir un ciclo de estiramiento y contracción forzados hasta que se hidrolizara todo el ATP agregado. En ese punto, la fibra nuevamente ya no se contraería..., o si se contrae, ya no podría estirarse.
La paradoja de la contracción fue que se requirió hidrólisis de ATP para la contracción muscular así como para la relajación (estiramiento). La paradoja se resolvió cuando finalmente se entendieron las funciones de los actores moleculares en contracción. Aquí revisamos algunos de los experimentos clásicos que llevaron a esta comprensión
4. Interacciones Actina-Miosina In Vitro: Disecciones y Reconstituciones
Un experimento temprano insinuó la interacción de actina y miosina en la contracción. Los homogeneizados de músculo esquelético fueron viscosos. Se aisló el componente viscoso y se demostró que contenía una sustancia que se denominó actomiosina (acto, activo; miosina, sustancia muscular). En condiciones apropiadas, la adición de ATP a las preparaciones de actomiosina provocó una disminución de la viscosidad. Sin embargo, después de hidrolizar el ATP agregado, la mezcla volvió a ser viscosa. La extracción de la preparación no viscosa (antes de volver a congelarse y antes de que se consumiera el ATP) condujo a la separación bioquímica de dos de las principales sustancias que ahora reconocemos como los filamentos de contracción de actina y miosina. Además, la adición de estos componentes de nuevo juntos reconstituyó el extracto viscoso de actomiosina (ahora denominado actinomiosina para reflejar su composición). Y..., agregar ATP a la solución reconstituida eliminó su viscosidad. Los cambios de viscosidad dependientes de ATP de las soluciones de actinomiosina fueron consistentes con una separación dependiente de ATP de filamentos gruesos y delgados. Quizás la actina y la miosina también se separan en los músculos glicerinados expuestos al ATP, lo que les permite estirarse y relajarse.
El advenimiento de la microscopía electrónica proporcionó más evidencia de un papel del ATP tanto en la contracción como en la relajación del músculo esquelético La purificación de la actina del músculo esquelético (todavía unida a las Líneas Z) a partir de la miosina se dibuja a continuación, mostrando cómo se veían los componentes separados en el microscopio electrónico.
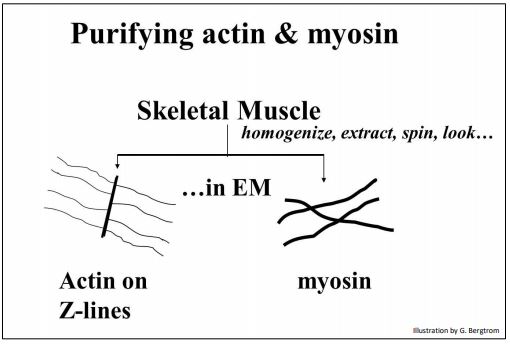
A continuación, cuando se mezclaron actina (todavía unida a las líneas Z) y miosina, la microscopía electrónica del material viscoso resultante reveló filamentos delgados que se entrelazaban con filamentos gruesos. A continuación se muestra el resultado de este experimento de reconstitución.
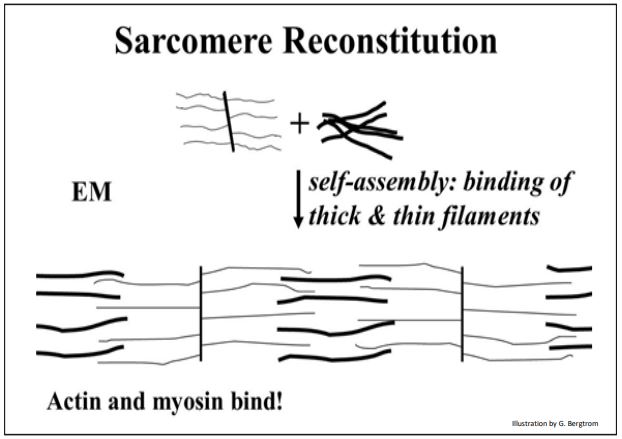
Como era de esperar, cuando se agregó ATP a estos extractos, la viscosidad de la solución disminuyó, y la microscopía electrónica que los filamentos gruesos revelados (miosina) y delgados (actina) se habían separado nuevamente. Los dos componentes pudieron aislarse de nuevo y separarse por centrifugación.
En experimentos adicionales, las preparaciones de actinomiosina podrían extenderse sobre una superficie acuosa, formando una película sobre la superficie del agua. Cuando se agregó ATP al agua, la película se “contrajo” visiblemente, alejándose de los bordes del recipiente, ¡reduciendo su superficie! La microscopía electrónica de la película reveló estructuras acortadas similares a sarcómeros con líneas Z estrechamente espaciadas y bandas I cortas..., confirmando aún más el modelo de filamento deslizante de contracción muscular.
332 Evidencia de microscopios in vitro y electrónicos para un modelo de filamento deslizante
Cuando la actina y la miosina se purificaron adicionalmente a partir de actinomiosina aislada, las barras gruesas de miosina pudieron disociarse en monómeros grandes de miosina de ~599Kd. Así, ¡los filamentos gruesos son polímeros masivos de monómeros de miosina! A continuación se muestra la estructura molecular de los filamentos gruesos de miosina.
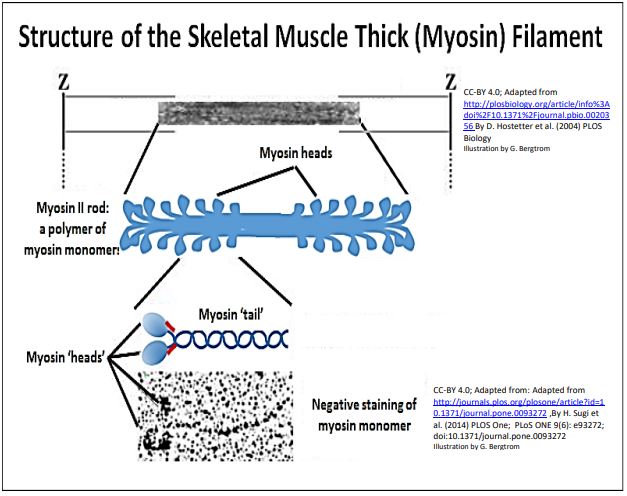
Una observación temprana de filamentos de actina aislados fue que no tenían actividad ATPasa. Por otro lado, mientras que las preparaciones de miosina aisladas sí tenían actividad ATPasa, solo catalizarían la hidrólisis de ATP muy lentamente en comparación con las fibras musculares intactas. La hidrólisis de ATP más rápida se produjo solo si los filamentos de miosina se mezclaron con microfilamentos (ya sea sobre, o desprendidos de las líneas Z). En el microscopio electrónico, los monómeros aislados de proteína miosina parecían tener regiones de doble cabeza y cola única. El análisis bioquímico mostró que los propios monómeros de miosina estaban compuestos por los dos polipéptidos de cadena pesada y dos pares de polipéptidos de cadena ligera mostrados en la ilustración anterior
Micrografías electrónicas de alto aumento, alta resolución y la ilustración correspondiente a continuación muestran las estructuras componentes de los monómeros de miosina.
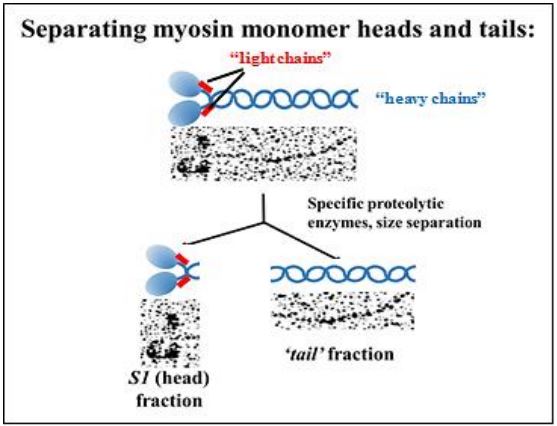
Las enzimas proteolíticas que hidrolizan enlaces peptídicos solo entre aminoácidos específicos, pueden 'cortar' las cadenas pesadas de los monómeros de miosina en fragmentos S1 (cabeza) y cola. Las micrografías electrónicas de estos dos fragmentos después de la separación por ultracentrifugación se muestran arriba. Se demostró que los fragmentos S1 tenían una actividad ATPasa lenta, mientras que las colas no tuvieron ninguna. La actividad lenta no fue un artefacto de aislamiento; la mezcla de la fracción S1 con filamentos de actina aislados resultó en una mayor tasa de hidrólisis de ATP. Claramente, las cabezas de miosina son ATPasas que interactúan con microfilamentos de actina.
Estructura de monómero de miosina y filamento grueso 333
A continuación se muestra la demostración directa de una asociación de fragmentos de cabeza de miosina S1 con microfilamentos de actina de músculo liso de conejo.

Al igual que para el músculo esquelético, la contracción del músculo liso se debe al deslizamiento de actina-miosina, aunque el músculo liso no está estriado y carece de morfología de sarcómero; una flecha blanca en la micrografía apunta a uno de varios filamentos de miosina (gruesos) visibles en la micrografía. La interacción de las cabezas de miosina S1 con filamentos de actina altera drásticamente su morfología. En esta imagen, las franjas diagonales, o la apariencia similar a la punta de flecha de la unión de S1-actina a lo largo de los filamentos de actina indica que los filamentos de F-actina son polares, con un extremo más (+) y un extremo menos (—), como se esperaba.
La misma “decoración” de microfilamentos con puntas de flecha se observa cuando las cabezas S1 (o incluso monómeros de miosina intactos) se unen a secciones delgadas de sarcómeros del músculo esquelético, preparaciones de actina aún unidas a las líneas Z, y con preparaciones aisladas de Factina. Estas imágenes son consistentes con el requisito de que la miosina debe unirse a la actina para lograr una tasa máxima de actividad de ATPasa durante la contracción.
Las puntas de flecha de la actina decorada aún unidas a las líneas Z siempre miran en direcciones opuestas, como se muestra a continuación.
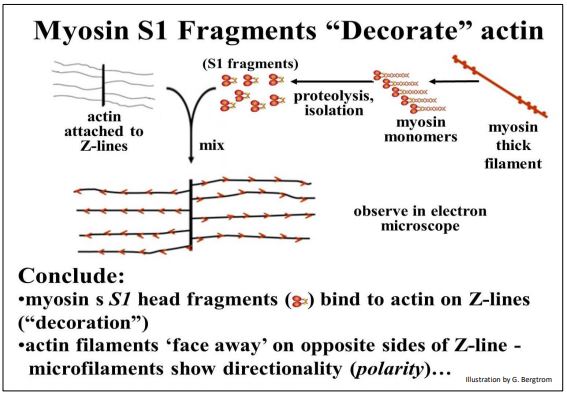
Estas puntas de flecha opuestas, consistentes con el modelo de contracción de filamentos deslizantes en el que el filamento grueso bipolar tira de los filamentos de actina uno hacia el otro desde lados opuestos de las varillas de miosina, acercando las líneas Z y acortando los sarcómeros.
5. Cambio alostérico y el ciclo de microcontracción
Mientras que la dineína y la cinesina son proteínas motoras que 'caminan' por los microtúbulos, el monómero de miosina es una proteína motora que camina a lo largo de los microfilamentos En cada caso, estas proteínas motoras son ATPasas que utilizan energía libre de hidrólisis de ATP para efectuar cambios conformacionales que resultan en la marcha, es decir, motilidad. En el músculo esquelético, los cambios alostéricos en las cabezas de miosina permiten que las varillas de miosina caminen a lo largo de los filamentos de F-actina
Cuando se colocan en secuencia, tales conformaciones de cabeza de miosina diferentes son probablemente las mismas que ocurrirían durante un ciclo de microcontracción (ilustrado a continuación).
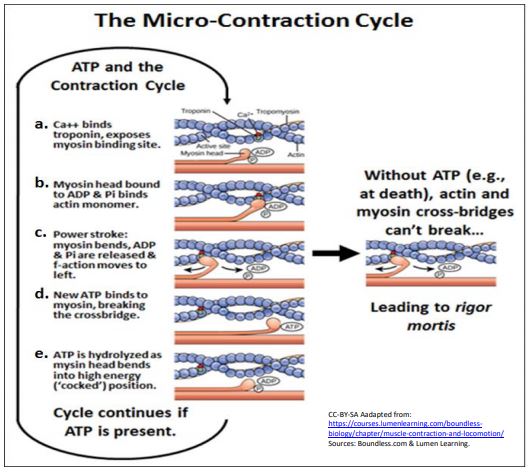
Para ayudarte a seguir la secuencia, sigue el pequeño punto rojo en un solo monómero en el filamento de actina. Estos son los pasos:
a. en presencia de iones Ca 2+, los sitios de unión a la miosina en la actina están abiertos (Ca 2+ - la regulación de la contracción muscular se discute con más detalle a continuación).
b. Las cabezas de miosina con ADP y Pi unidos se unen a sitios abiertos en filamentos de actina.
c. El resultado de la unión actina-miosina es un cambio alostérico en la cabeza de la miosina, una flexión de la región bisagra, que tira del microfilamento unido (siga el punto rojo - ¡se ha movido de derecha a izquierda!). Este poco de micro-deslizamiento de actina a lo largo de la miosina es el golpe de potencia.
d. En su conformación 'doblada', la cabeza de miosina, aún unida a un monómero de actina en la F-actina, se une al ATP, haciendo que ADP y Pi salgan de la cabeza de miosina y la disocien de la actina.
e. Una vez disociadas de la actina, las cabezas de miosina catalizan la hidrólisis de ATP, dando como resultado otro cambio conformacional. La cabeza, aún unida a ADP y Pi, se ha doblado en su bisagra, adquiriendo una conformación de alta energía que almacena la energía de la hidrólisis de ATP.
f. La energía libre almacenada se libera durante la carrera de potencia. Si se ha eliminado Ca 2+, las miosinas permanecen en la conformación de alta energía del paso e, hasta que una liberación de Ca 2+ nuevamente señala la contracción.
Los ciclos de microcontracción de actina que se desliza a lo largo de la miosina continúan mientras el ATP esté disponible. Durante ciclos repetitivos de microcontracción, las cabezas de miosina en los filamentos gruesos tiran de los filamentos de actina unidos a las líneas Z, acercando las líneas Z entre sí. El resultado es acortamiento del sarcómero y en última instancia, de las células musculares y de todo el músculo. En ausencia de ATP (como después de la muerte de un organismo), se interrumpe el ciclo de microcontracción. Todas las cabezas de miosina permanecerán unidas a los filamentos de actina en estado de contracción o relajación muscular (estiramiento) al momento de la muerte. Se trata de rigor mortis a nivel molecular (ver la ilustración anterior). A nivel de todo el músculo, el rigor mortis resulta en la incapacidad de estirar o de otra manera mover partes del cuerpo cuando el ATP está, de una vez por todas, agotado.
6. Resolviendo la paradoja de la contracción
El ciclo de microcontracción de la cabeza de miosina resuelve la paradoja de la contracción:
- El ATP es necesario para la contracción muscular: En el paso e de la ilustración anterior, a medida que el ATP en las cabezas de miosina se hidroliza, las cabezas cambian de una conformación de baja energía a una de alta energía. Las cabezas de miosina ahora pueden unirse a monómeros de actina (etapa b en el ciclo de microcontracción). Esto resulta en el golpe de potencia (paso c), donde la energía libre liberada por un cambio alostérico en la miosina, tira de la actina a lo largo de la miosina, en efecto provocando un micro-acortamiento del sarcómero, en otras palabras, ¡contracción!
- El ATP es necesario para la relajación muscular: Al final del paso c, la miosina permanece unida a la actina hasta que el ATP puede unirse nuevamente a la cabeza de la miosina. La unión de ATP en la etapa d desplaza el ADP y el fosfato inorgánico (Pi) y rompe los puentes cruzados de actinmiosina. Una eliminación de Ca 2+ de los sarcómeros al final de un evento de contracción bloquea los sitios de unión de la miosina en la actina, mientras que la rápida rotura de los puentes cruzados de actina y miosina por la unión de ATP-miosina permite la relajación muscular y el deslizamiento de los filamentos de actina y miosina (es decir, estiramiento). Esto deja a las cabezas de miosina en la conformación 'ameteada' (alta energía), listas para la siguiente ronda de contracción.
En resumen, la unión a ATP-miosina rompe los puentes cruzados actina-miosina. El músculo puede entonces relajarse y estirarse. La energía libre de la hidrólisis de ATP, ahora almacenada en una conformación de miosina de alta energía, se libera durante la carrera de potencia de microcontracción
El examen microscópico electrónico de las cabezas de monómero de miosina a diferentes fuerzas iónicas o cuando se unen a anticuerpos (como se muestra a continuación), proporciona evidencia visual de que las cabezas de miosina son flexibles y pueden tomar conformaciones estables alternas, como se esperaría durante el ciclo de microcontracción.
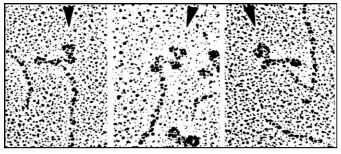
Las puntas de las flechas apuntan a moléculas de anticuerpo unidas (inmunoglobulinas). Para un video de cambio conformacional en monómeros de miosina en cabezas de miosina en acción.
335 Un ciclo de contracción de actina-miosina resuelve la paradoja de la contracción
336 La unión e hidrólisis de ATP cambia la conformación de la cabeza de miosina
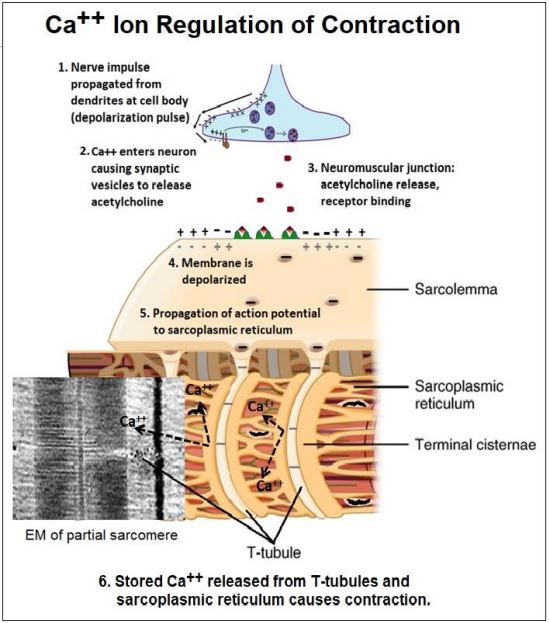
7. Los iones Ca 2+ regulan la contracción del músculo esquelético
Típicamente, el neurotransmisor acetilcolina liberado por una neurona motora se une a receptores en las células musculares para iniciar la contracción. En los primeros experimentos, se requirió Ca 2+, junto con ATP, para que el músculo esquelético glicerinado se contrajera. Posteriormente se demostró que los iones Ca 2+ se almacenaron en el retículo sarcoplásmico, el retículo endoplásmico liso de las células musculares. Como hemos visto, un potencial de acción generado en el cuerpo celular de una neurona se propaga a lo largo de un axón hasta el terminal nervioso, o sinapsis. De manera similar, un potencial de acción generado en una unión neuromuscular viaja a lo largo del sarcolema (la membrana plasmática muscular) hasta puntos donde es continuo con túbulos transversales (túbulos T). El potencial de acción luego se mueve a lo largo de los túbulos T y luego a lo largo de las membranas del retículo sarcoplásmico. Esta propagación de un potencial de acción abre canales de Ca 2+ en el retículo sarcoplásmico. El Ca 2+ liberado baña los sarcómeros de las miofibrillas, permitiendo que los filamentos se deslicen (es decir, contracción). El potencial de acción en una unión neuromuscular que inicia la contracción se resume en la siguiente ilustración.
Los iones Ca 2+ liberados del retículo sarcoplásmico bañan las miofibrillas, donde se unen a una de las tres moléculas de troponina para regular la contracción del músculo esquelético. Las tres troponinas y una molécula de tropomiosina están unidas a filamentos de actina. Los experimentos con anticuerpos anti-troponina y anti-tropomiosina localizan estas proteínas en filamentos delgados espaciados a intervalos regulares en micrografías electrónicas. El siguiente dibujo modela esta asociación de las subunidades de troponina y tropomiosina con los filamentos delgados.
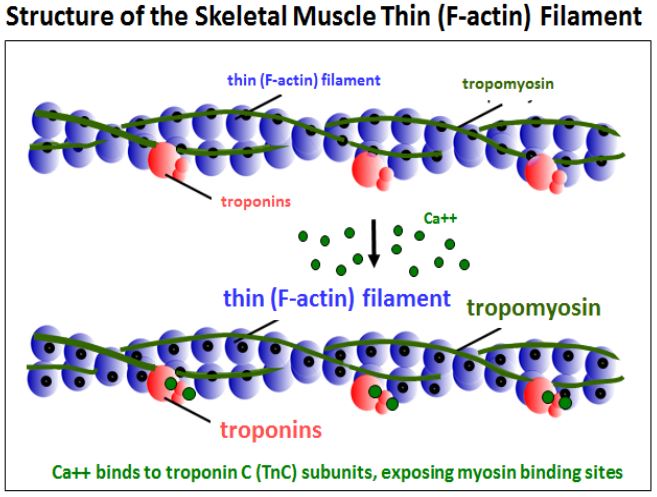
En el músculo en reposo, la tropomiosina (una proteína fibrosa) se encuentra a lo largo del filamento de actina donde cubre los sitios de unión a la miosina de siete subunidades de G-actina en el microfilamento. En esta conformación, la troponina T (troponina de unión a tropomiosina) y la troponina I (troponina inhibitoria) mantienen la tropomiosina en su lugar. La siguiente ilustración de sección transversal ilustra el papel de los cambios conformacionales en la troponina C en la unión de Ca++ en la regulación de la contracción.
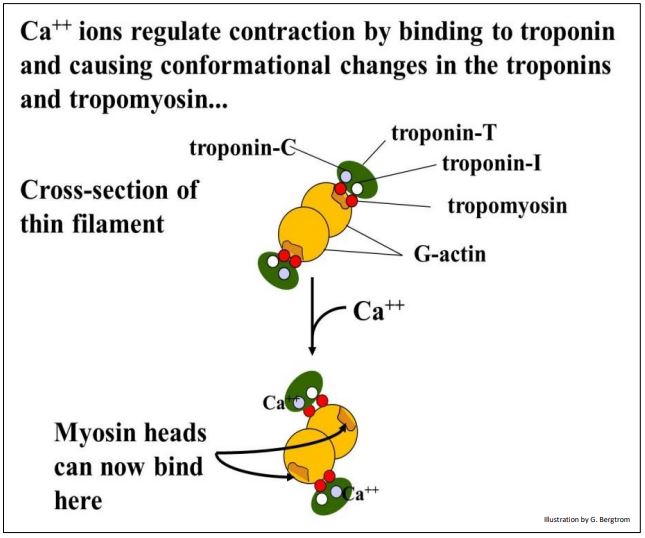
337 Regulación de la contracción del músculo esquelético por calcio
8. La contracción muscular genera fuerza
La contracción por deslizamiento impulsado por ATP de filamentos delgados a lo largo de gruesos genera fuerza en las líneas Z. En tres dimensiones, las líneas Z son en realidad discos Z) a los que se unen los filamentos delgados de actina. La \(\alpha \)proteína-actinina en los discos Z ancla los extremos de los filamentos de actina a los discos de manera que cuando los filamentos se deslizan, los discos Z se acercan, acortando los sarcómeros. Otra proteína del disco Z, la desmina, es un filamento intermedio organizado alrededor de la periferia de los discos Z. Desmin conecta múltiples discos Z en una miofibrila. Al mantener los Z-Disks en registro, se coordina la célula muscular y, en última instancia, la contracción muscular. Finalmente, los filamentos de actina en los extremos de la célula deben conectarse a la membrana celular para que una célula muscular se acorte durante la contracción de la miofibrilla. Varias proteínas, entre ellas las sintrofinas y la distrofina (otra proteína de filamento intermedio) anclan los extremos libres de los microfilamentos provenientes de los discos Z a la membrana celular. Aún otras proteínas anclan la membrana celular en esta región a la matriz extracelular (tendones) que a su vez están, ¡unidas a los huesos! La fuerza generada por la hidrólisis de la miosina del ATP y el deslizamiento de filamentos en sarcómeros individuales se transmiten así a los extremos de los músculos para efectuar el movimiento. Si el nombre distrofina suena familiar, ¡debería! El gen y su proteína fueron nombrados por una mutación que causa distrofia muscular, resultando en un debilitamiento muscular progresivo.
338 Contracción Genera Fuerza Contra Discos Z y Membranas Celulares
9. El Sarcómero Elástico: ¿Las Varillas de Miosina Simplemente Flotan en el Sarcómero?
De hecho, las barras de miosina están ancladas a proteínas en los discos Z y líneas M. En 1954, R. Natori se dio cuenta de que cuando el músculo contraído se relaja, se alarga más allá de su estado de reposo, para luego acortarse nuevamente a su longitud de reposo. Propuso que esta elasticidad debe deberse a una fibra en el sarcómero. Veinticinco años después, la estructura elástica fue identificada como titina, ¡una proteína que contiene varios registros moleculares! El gen de la titina contiene el mayor número de exones (363) de proteínas conocidas. Después de la actina y la miosina, la titina es también la proteína más abundante en las células musculares. Con casi 4 x 106 Da, el acertadamente llamado titina es también el polipéptido más grande conocido. Extendiéndose desde los discos Z hasta la línea M de sarcómeros, el titín se enrolla alrededor de filamentos gruesos a lo largo del camino. La titina está anclada en los discos Z por las proteínas \(\alpha \)-actinina y teletonina. En la línea M, la titina se une a la proteína C de unión a miosina (MYBPC3) y calmodulina, entre otras (por ejemplo, miomesina, obscurin y esquelamina). Algunas, si no todas, estas proteínas deben participar en mantener los filamentos gruesos de miosina posicionados y registrados en el sarcómero. Esto es similar a cómo los discos Z unen los extremos de los filamentos de actina para mantener los sarcómeros registrados. La ubicación de la titina y varias otras proteínas de sarcómero se ilustra a continuación.
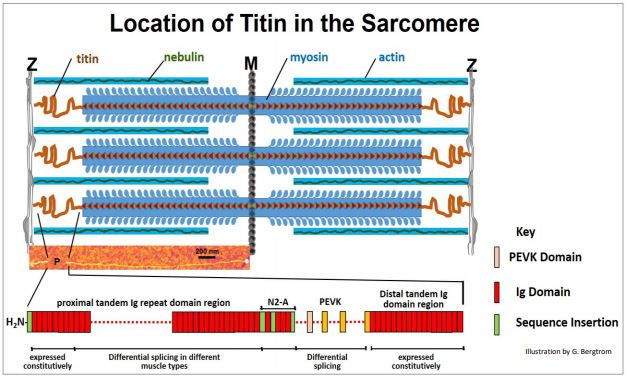
Las moléculas de titina enrolladas (en rojo en la ilustración) se extienden desde las líneas Z a M. La micrografía electrónica coloreada de una molécula de titina extendida en el medio de la ilustración anterior debería convencerte de la longitud (¡35,213 aminoácidos!) de este enorme polipéptido! Las características elásticas de Titín se encuentran principalmente en la región etiquetada P en la micrografía, entre los discos Z y las varillas de miosina. Los muchos dominios de esta región P se muestran en la parte inferior de la ilustración. Con todas las funciones de enlace (y otras), puede esperar que titin tenga muchos dominios. ¡Lo hace! Incluyen dominios de Ig (inmunoglobulina), dominios de fibronectina (no mostrados aquí), dominios PEVK y N2A (que ayudan a unir la titina a\(\alpha \) -actinina en los discos Z). Cuál y cuántos dominios Ig y/o PEVK están presentes en un músculo particular depende de qué vía alternativa de corte y empalme se use para formar un ARNm de titina. Más de una micra de largo, Titín funciona como un resorte molecular, como predijo Natori. Sus dominios enrollados se comprimen durante la contracción, almacenando pasivamente parte de la energía de contracción. Cuando el músculo esquelético se relaja, el Ca 2+ se retira del sarcómero, el ATP desplaza el ADP de las cabezas de miosina y la actina y la miosina se disocian. El músculo luego se estira, típicamente bajo la influencia de la gravedad o un conjunto opuesto de músculos. Sin embargo, durante la contracción, 244 dominios proteicos plegados individualmente de la titina se comprimieron, y durante la relajación, estos dominios se descomprimen; la energía almacenada de compresión también ayuda a potenciar la relajación. Al mismo tiempo, las conexiones de titina limitan el estiramiento para que un músculo potencialmente sobreestirado pueda 'rebotar' a su longitud relajada normal.
En un experimento particularmente elegante, R. Linke et al. proporcionaron una demostración visual de la elasticidad de miofibras consistente con el modelo de resorte enrollado de la estructura de la titina. Hicieron anticuerpos contra dominios peptídicos a ambos lados del dominio PEVK de la titina (N2A e I20-I22) y los unieron a partículas nanogoladas (que aparecerán como gránulos densos de electrones en microscopía electrónica de transmisión). Luego, las miofibras individuales se estiraron a diferentes longitudes, se fijaron para microscopía electrónica y se trataron con los anticuerpos ligados a nanogold. Los anticuerpos localizan y definen los límites de los dominios de la titina PEVK. La imagen de abajo no muestra micrografías electrónicas originales teñidas inmunológicamente, pero muestra micrografías alternas de sarcómeros con localización simulada de partículas nanogold, reflejando resultados reales.
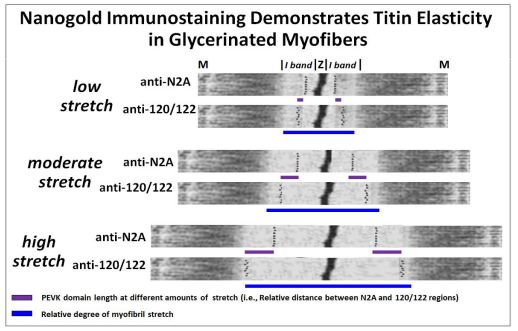
En el experimento, el aumento del estiramiento alargó las bandas I a ambos lados de las líneas Z de sarcómeros (barras azules). Asimismo, los dominios PEVK de titina también se alargaron como es evidente a partir de la mayor distancia entre los anticuerpos N2A y 120/122 unidos a nanogold que se unen a ambos lados de los dominios PEVK. Esta demostración de la elasticidad de la titina (y por lo tanto del sarcómero) es consistente con el almacenamiento de parte de la energía libre de contracción cuando la molécula se comprime, y la liberación pasiva de esa energía durante la relajación. Dado que la titin ata filamentos gruesos a Zdisks y líneas M, también limita la cantidad de estiramiento del sarcómero durante la relajación. Una animación del laboratorio de Linke se encuentra en http://www.titin.info/.
D. Microfilamentos no musculares
La microscopía electrónica reveló que filamentos delgados (~10 nm) permearon el citoesqueleto de las células eucariotas. Se sospechó que eran microfilamentos de actina. Los microfilamentos generalmente se encuentran en la corteza de las células, justo debajo de la membrana plasmática, donde soportan la forma celular. Estos mismos microfilamentos también pueden reorganizarse dinámicamente, permitiendo que las células cambien de forma. Un ejemplo dramático de esto ocurre en las células en división, durante la citocinesis cuando la célula en división forma un surco de escisión en el medio de la célula. Los microfilamentos corticales se deslizan uno junto al otro con la ayuda de la miosina no muscular, pellizcando progresivamente la célula hasta que se divide en dos nuevas células.
Para probar si estos 'microfilamentos' de 10 nm eran de hecho actina, monómeros de miosina intactos o fragmentos de cabeza de miosina S1 se colocaron sobre micrografías electrónicas de muchos tipos diferentes de células. Al verse en el microscopio electrónico, tales preparaciones siempre revelaron que los microfilamentos de 10 nm estaban decorados con puntas de flecha, ¡al igual que la actina de células musculares decorada con fragmentos S1 o la actina unida a la línea Z! Claramente, estos microfilamentos citoplásmicos son una forma de F-actina. En el ejemplo que se muestra a continuación, las células en citocinesis se trataron con fragmentos de cabeza de miosina S1.
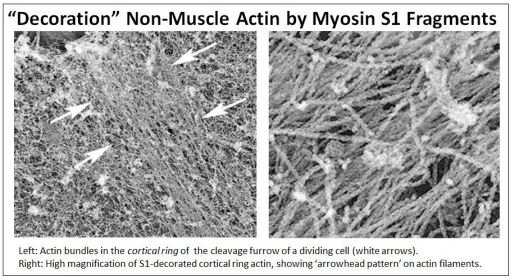
Vea el papel de los filamentos corticales en la citocinesis en la Acción del Filamento de Actina Cortical en la Citocinesis. Por supuesto, los microfilamentos de actina están involucrados en todo tipo de motilidad celular además de su papel en la división celular. Permiten el movimiento celular y el flujo citoplásmico dentro de las células. Y aunque dan fuerza a las microvellosidades intestinales, incluso permiten que las microvellosidades se muevan independientemente de las presiones pasivas del peristaltismo.
Otros ejemplos de microfilamentos en la motilidad celular incluyen la capacidad de la ameba y otras células fagocíticas para extender los pseudopodios para engullir alimentos o partículas extrañas (por ejemplo, bacterias), respectivamente. De manera similar, cuando las células de fibroblastos se mueven a lo largo de las superficies, extienden filipodios delgados en la dirección del movimiento ensamblando haces de actina a lo largo del eje de movimiento celular. Fibras de estrés de actina que ayudan a mantener la forma celular de fluorescencia verde en la micrografía de inmunofluorescencia a continuación (panel izquierdo). También se ilustran los papeles duales de la actina en el movimiento de los fibroblastos (abajo a la derecha).
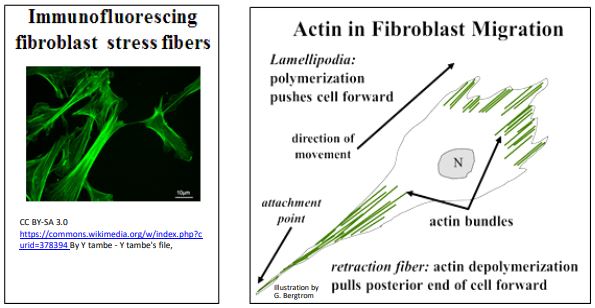
Como vimos para la motilidad celular mediada por microtúbulos, alguna motilidad mediada por actina puede estar basada principalmente en el ensamblaje y desmontaje de actina, como en la extensión de filipodios en el frente móvil de un fibroblasto. A medida que el fibroblasto avanza, una fibra de retracción en el extremo posterior de la célula permanece unida a la superficie (sustrato) a lo largo de la cual está migrando. Finalmente, sin embargo, las interacciones actina-miosina (de hecho, deslizamiento) provocan la retracción de la mayor parte de esta 'fibra' de nuevo en el cuerpo de la célula.
Los movimientos mediados por fibras de estrés también pueden explicar el flujo citoplásmico que distribuye los componentes celulares y nutrientes a través de una célula. Los movimientos de ambos implican interacciones actina-miosina. Los estudios de motilidad de células no musculares sugieren la estructura y los componentes moleculares que interactúan de las fibras de estrés. Revelan filamentos superpuestos de miosina y actina que se deslizan durante el movimiento, como se ilustra a continuación.
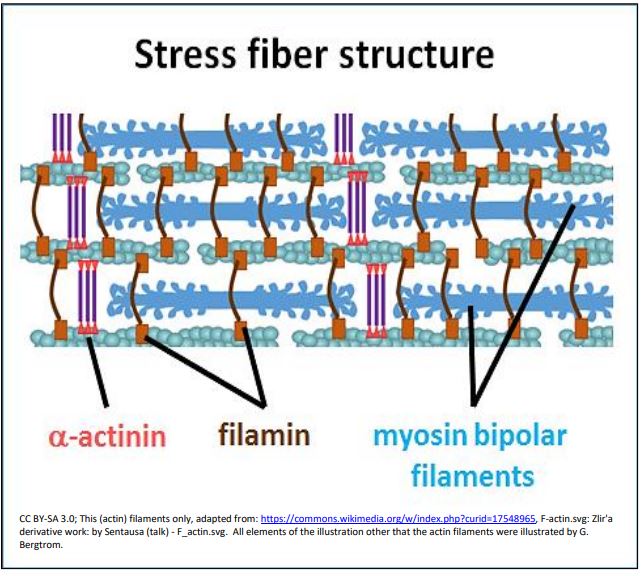
La filamina en este dibujo se muestra manteniendo unidos los filamentos de actina en ángulo, mientras que\(\alpha \) la actinina también ayuda a agrupar los filamentos de actina (delgados). La titina (no mostrada) también parece estar asociada con fibras de tensión. Sin embargo, a diferencia de los sarcómeros altamente organizados del músculo esquelético, las proteínas y filamentos en las fibras de estrés no forman parte de las superestructuras de la línea Z o M. ¿Podrían esos haces de filamentos de fibra de estrés no muscular menos organizados ser el predecesor evolutivo de los sarcómeros en las células musculares?
E. Actinas y miosinas son codificadas por grandes familias génicas
¡Las actinas pueden ser la proteína más abundante en las células! Al menos seis diferentes isoformas de actina codificadas por una gran familia de genes de actina tienen secuencias de aminoácidos casi idénticas, todas las cuales están involucradas en la función citoesquelética. Predomina la isoforma\(\beta \) -actina. Los genes para algunas isoformas se expresan de una manera específica de la célula.
¿Todas las isoformas de actina son funcionalmente significativas? Los monómeros de miosina (o cabezas S1) decoran prácticamente todas las actinas. Esto hace que uno se pregunte si alguna actina es una adaptación, por sutil que sea, tal que la ausencia de una isoforma representaría una amenaza significativa para la supervivencia de un organismo. Dado que las diferencias de secuencia de aminoácidos entre actinas no predecirían una función proteica dramáticamente diferente, ¿podrían subyacer alguna ventaja fisiológica aún desconocida para diferentes células? En ratones, la pérdida del gen a\(\gamma \) -actina tiene poco efecto sobre el organismo, mientras que la pérdida del gen\(\beta \) -actina en ratones es letal en etapas embrionarias. En contraste, los estudios muestran que un gen mutado\(\beta \) -actina en humanos se correlaciona con retraso en el desarrollo y problemas neurológicos posteriores (por ejemplo, epilepsia), anomalías renales y cardíacas, pero no es letal. De hecho, las personas con tales mutaciones pueden llevar vidas casi normales y saludables (mutaciones y enfermedades del gen de la beta-actina).
Al igual que las actinas, los genes de miosina que codifican isoformas variantes comprenden una gran familia de genes eucariotas. Todas las isoformas tienen actividad ATPasa y algunas están claramente involucradas en la motilidad celular. Las funciones únicas aún no se conocen para otras isoformas, pero diferentes monómeros de miosina pueden decorar actina, y las miosinas de una especie pueden decorar filamentos de actina de otras especies, incluso a través de amplias distancias filogenéticas.
F. Filamentos intermedios - una visión general
Estos filamentos de 10 nm son proteínas con estructura secundaria extendida que de hecho, no se pliegan fácilmente en estructuras terciarias, y no tienen actividad enzimática. Recordemos su ubicación intercelular en los desmosomas donde unen firmemente las células para conferir resistencia a la tracción a los tejidos. Dentro de las células, los filamentos intermedios permean las células donde participan en la regulación y mantenimiento de la forma celular. Recordemos su papel en el anclaje de actina a discos Z o placas de membrana plasmática en las células musculares, transmitiendo las fuerzas de contracción al acortamiento de los sarcómeros y luego al acortamiento real de un músculo. Las queratinas extracelulares que componen el pelaje, el pelo, las uñas de las manos y los pies, son proteínas relacionadas con filamentos intermedios. A diferencia de los filamentos intermedios intracelulares, las queratinas son haces de proteínas extracelulares rígidas e insolubles que se combinan para alinearse para formar estructuras secundarias estables e inmutables. Finalmente, las laminas son filamentos intermedios que conforman elementos estructurales de la lámina nuclear, una especie de nucleoesqueleto.
CMB3e 492 Como vimos anteriormente, las subunidades de filamentos intermedios tienen una estructura común que consiste en un par de monómeros, cada uno con dominios globulares en sus extremos C y N-terminales, separados por regiones de varilla enrollada. Los monómeros son no polares; es decir, a diferencia de los microtúbulos y filamentos de actina, no tienen extremos 'más' y 'menos'. La unidad básica de estructura de filamento intermedio es un dímero de monómeros. Los dímeros se agregan más para formar tetrámeros y haces de filamentos más grandes. Al igual que los microtúbulos y los filamentos de actina, los haces de filamentos intermedios se pueden desmontar y volver a ensamblar según sea necesario cuando las células cambian A diferencia de los microtúbulos y la actina, los filamentos intermedios pueden estirarse, una propiedad conferida por las regiones de las varillas enrolladas de los filamentos. ¡Esto debería ser una reminiscencia de las moléculas de titina! Las características estructurales y la elasticidad de los filamentos intermedios se ilustran en la caricatura a continuación.
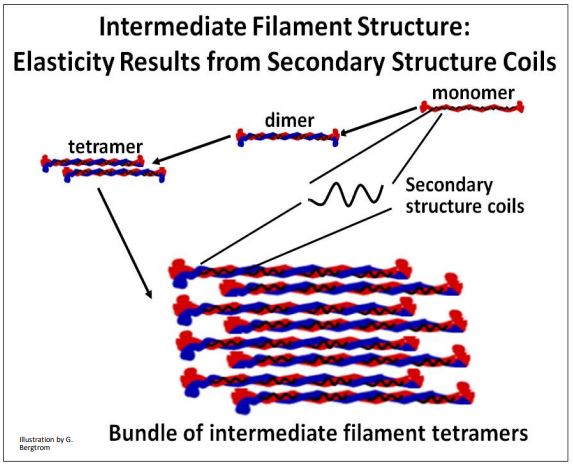
En los filamentos intermedios agrupados que permean el citoplasma de las células, la capacidad de estiramiento contribuye a la viscosidad del citoplasma, e incluso se denomina viscoelasticidad. Se cree que esta propiedad elástica permite a las actinas y microtúbulos un grado de libertad de movimiento de las células, y dentro del citoplasma de las células.