
19.4: Cuando las celdas mueren
- Page ID
- 54190

Como se señaló, pocos tipos de células viven para siempre; la mayoría vive por un tiempo finito. La mayoría están destinados a darse la vuelta (otro eufemismo para morir), mediada por muerte celular programada, o apoptosis. Esto ocurre en el desarrollo normal cuando las células solo se requieren temporalmente para un proceso de maduración (por ejemplo, desarrollo embrionario, metamorfosis). Cuando ya no es necesario o cuando se dañan genéticamente o de otra manera, dichas células son detectadas y señaladas para que sufran apoptosis. La muerte celular programada comienza con una señal externa programada para aparecer en un momento específico de desarrollo. La molécula señal actúa sobre las células diana para inducir la transcripción de genes Bcl2. Las proteínas Bcl2 Bak y Bax son componentes externos del canal de la membrana mitocondrial que permiten la liberación del citocromo C en el citoplasma. Esto desencadene eventos moleculares que conducen a la apoptosis. A continuación se ilustra el papel del citocromo C en la apoptosis.
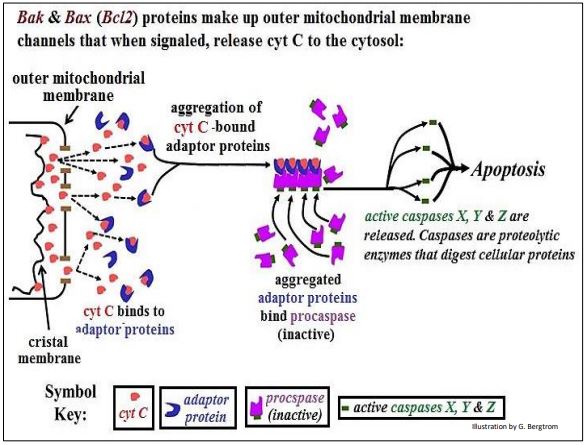
La salida mitocondrial del citocromo C es posible porque es una proteína de membrana periférica, solo ligada flojamente a la membrana de cristal. Existe en equilibrio entre los estados unidos a membrana y no unidos. A medida que algunas moléculas de citocromo C salen del espacio intermembrana, otras se desprenden de la membrana de cristal y siguen. En el citosol, el citocromo C se une a proteínas adaptadoras que luego se agregan. El complejo citocromo c-adaptador tiene una alta afinidad por una procaspasa biológicamente inactiva. La unión de procaspasa al complejo citocromo adaptador C provoca un cambio alostérico en la procaspasa, liberando una caspasa activa. Las caspasas son enzimas proteolíticas que inician la autodigestión de la célula.
Un ejemplo de apoptosis es la metamorfosis de anfibios. La hormona tiroidea señala metamorfosis de renacuajo. La hormona hace que los renacuajos digieran sus propias células de la cola, permitiendo la reabsorción y reciclaje de los productos de digestión. Estos a su vez sirven como nutrientes para cultivar estructuras de ranas adultas. Por su trabajo en la identificación de genes de apoptosis, Sydney Brenner, H. Robert Horvitz y John E. Sulston compartieron el Premio Nobel de Fisiología o Medicina 2002.