
1: Anatomía de una célula - Una descripción muy breve
- Page ID
- 53308

Dado que todo este curso está dedicado a comprender el funcionamiento de la célula, resulta casi superfluo dedicar un capítulo a identificar las partes de la célula y sus funciones. Sin embargo, debido a que es fácil perderse en las complejidades de las moléculas y las reacciones químicas dentro de la célula, considere este capítulo más como un marco o mapa para el curso, dando contexto a las minucias.
La célula es la unidad de vida más pequeña, por lo que todas las células, ya sean organismos unicelulares o solo una pequeña parte de un organismo multicelular, tienen ciertas características en común: deben contener información genética y los mecanismos para regular y utilizar esa información para producir sus propias partes y para reproducir nuevas células, deben ser capaces de usar energía en reacciones químicas y acciones físicas, deben poder regular esas actividades, y deben responder a estímulos.
Las células usan ADN (ácido desoxirribonucleico) para su material genético, y todas las células contienen las enzimas transcripcionales y traduccionales para leerlo y usar la información para construir más componentes celulares. Sin embargo, el simple hecho de tener material genético no define la vida: los virus tienen material genético que contiene toda la información necesaria para hacer un virus completo, pero no contiene las enzimas necesarias para hacerlo, ni la capacidad de obtener la materia molecular prima necesaria para hacerlo. Es absolutamente dependiente de la maquinaria dentro de cualquier célula que infecta. Por lo tanto, un virus no es un organismo vivo.
El genoma no es sólo un conjunto de instrucciones para hacer una célula (o un organismo, para el caso); también es replicable en sí mismo. En términos generales, durante parte de su ciclo de vida, la célula hace una copia extra de su genoma y aumenta los números de todas las demás “cosas” (proteínas, grasas, etc.) de las que está hecha, y luego se reproduce por división. La célula madre se divide en dos células hijas, cada una con el mismo complemento de información genética y con aproximadamente los mismos componentes celulares. Así vemos que si bien el genoma a menudo se considera el modelo para una célula/organismo, de hecho las células no se construyen desde cero directamente a partir del ADN. Cada celda viene de otra celda. El ADN puede entonces ser utilizado para personalizar esa célula para fines específicos según lo determinado por su entorno. Cuando se necesita un componente particular de la célula, la información para hacer ese componente se lee del ADN y se copia en ARN que se usa como un programa a partir del cual los ribosomas pueden fabricar las proteínas necesarias. Una célula viva necesita todas estas cosas: la información genética, los mecanismos y maquinaria para usar la información para construir partes celulares, y la capacidad de aprovechar la energía para hacerlo.
Como veremos en el capítulo 3, las leyes físicas de la naturaleza requieren que todo tienda hacia su estado más simple, menos organizado, a menos que haya un insumo de energía para trabajar contra esa tendencia. Dado que las células son una colección altamente ordenada de moléculas muy complejas, por lo tanto deben requerir energía para permanecer como células. Así, la vida requiere la capacidad de obtener energía, ya sea de la luz solar o de los alimentos, y la capacidad de convertir esa energía en formas que puedan ser fácilmente utilizadas por la célula para mantenerse construyendo o reordenando las moléculas y estructuras macromoleculares necesarias.
¿Cómo saben las células cuándo llevar a cabo estas actividades? Esto nos lleva a la siguiente característica de las células vivas: la capacidad de responder a estímulos. En otras palabras, son autorregulables. Si los niveles de glucosa se reducen y la célula necesita energía, se elaboran proteínas de transporte de glucosa, o si la célula necesita moverse a un área de mayor concentración de alimentos, el citoesqueleto celular se reorganiza para mover la célula. La célula tiene la capacidad de iniciar procesos de reparación si detecta lesiones en su genoma, puede pausar el ciclo celular para permitir que dichos procesos de reparación tengan tiempo, e incluso puede iniciar su propia muerte si las reparaciones son repetidamente infructuosas.
Además de responder a las señales internas, las células vivas también son capaces de responder a estímulos externos. Ya sea por contacto con una célula vecina, uniéndose a una hormona liberada de una célula lejana, o simplemente interactuando con objetos ambientales no celulares, una célula es capaz de responder a tales estímulos. Las respuestas pueden incluir hacer nuevas proteínas, destruir las proteínas existentes, alejarse del estímulo, avanzar hacia el estímulo, iniciar la reproducción y muchas otras posibilidades.
Existen dos tipos básicos de células: procariotas y eucariotas. La diferencia es simple y fácilmente reconocible bajo microscopía óptica. Las células eucariotas contienen compartimentos intracelulares unidos a la membrana (llamados orgánulos). Las células procariotas no contienen ninguno de esos compartimentos (Figura\(\PageIndex{1}\)).
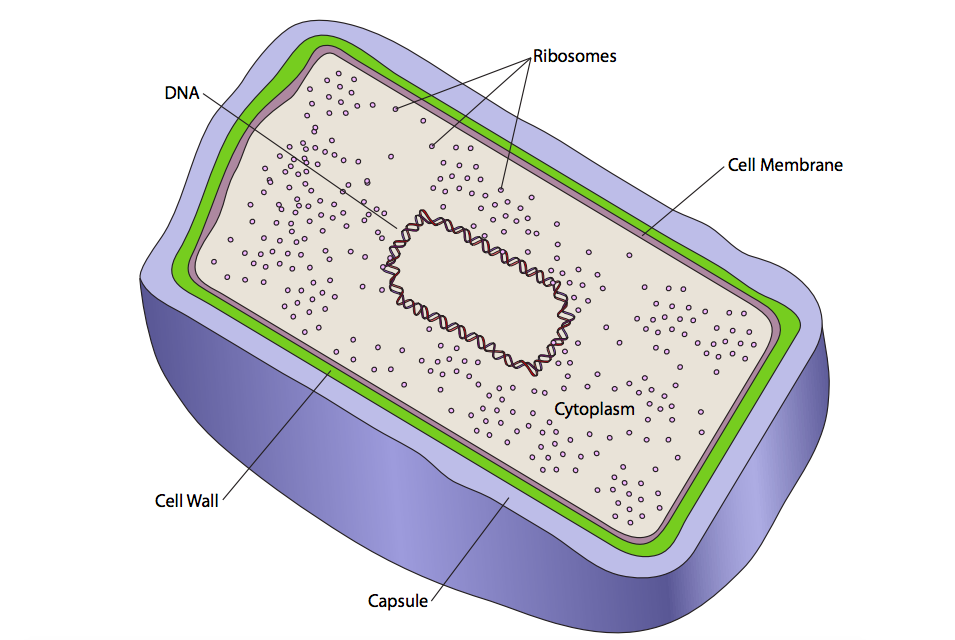
Solo hay una membrana en los procariotas, la membrana celular, y solo un compartimento en las células procariotas, el citoplasma. Eso no excluye cierto nivel de organización en los procariotas, pero no es tan complejo como los eucariotas. El ADN genómico suele estar organizado en un nucleoide central. No hay orgánulos membranosos intracelulares, pero la célula está definida por una membrana celular. Fuera de la membrana celular, los procariotas tienen una pared celular. Esta pared es relativamente rígida y confiere forma a la celda. Dependiendo del tipo de bacteria, el grosor de la pared varía (grueso = “gram positivo”, delgado = “gram negativo”). Algunas, pero no todas las bacterias también secretan otra capa fuera de la pared celular. Esta es una matriz relativamente apretada llamada cápsula que ayuda a proteger la célula de la desecación en ambientes secos. Una matriz comparativamente suelta de los mismos tipos de moléculas puede ser secretada, y en lugar de la cápsula, el resultado se llama capa de limo. La capa de limo es importante en la unión bacteriana y la formación de biopelículas (ver capítulo 13, Matriz extracelular).
Las células eucariotas son considerablemente más complejas. Actualmente, los organismos eucariotas se clasifican en cuatro reinos: animal, vegetal, hongo y protistas. La célula animal en la Figura\(\PageIndex{2}\) representa tiene muchas características en común con las células de los otros tres reinos.
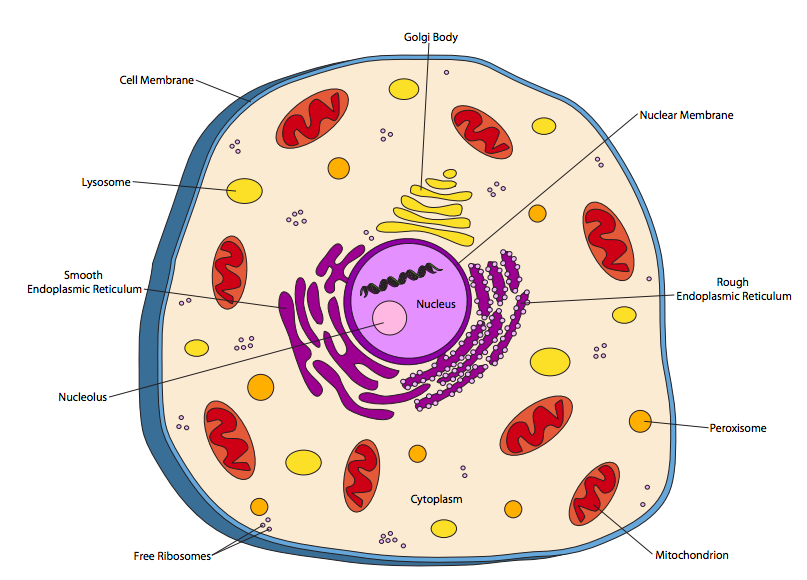
Obviamente, la mayor diferencia entre la célula animal (o cualquier célula eucariota) y las células procariotas es la presencia de compartimentos internos unidos a la membrana, u orgánulos. El más destacado de estos es el núcleo, que alberga el ADN. Tradicionalmente, se ha asumido que la mayoría de los genomas eucariotas pueden variar de 10 a 100 x 10 6 nucleótidos (10-100 Mb) de longitud total, sobre dos o más cromosomas (moléculas de ADN) de tamaño aproximadamente similar. Por el contrario, los genomas procariotas se han visto tradicionalmente como un solo cromosoma circular, y en su mayoría bajo una megabase (106 nucleótidos) de longitud.
Los métodos recientes y mejores para el mapeo y secuenciación del genoma, y la ampliación de los organismos de la muestra han demostrado que esos números son inexactos. De hecho, los genomas eucariotas oscilan entre ~3 Mb y más de 4000 Mb. Los genomas procariotas varían de 0.5 Mb a poco más de 10 Mb (0.5 a 6 Mb para Archaea, 0.6 a 10 Mb para Bacterias) y pueden extenderse sobre múltiples moléculas de ADN que pueden ser lineales o circulares.
El núcleo está limitado por una membrana de doble capa (la mayoría de los otros orgánulos están limitados por una sola membrana) que es continua con el retículo endoplásmico (ER). El retículo endoplásmico se subdivide en el RE rugoso (RER) y el RE liso (SER) en base a la aparición en micrografías electrónicas. Los “sementales” en el RER son ribosomas, que son la maquinaria molecular para elaborar proteínas en la célula. También hay ribosomas de flotación libre -la diferencia es que los ribosomas libres producen proteínas que permanecen en el citoplasma, mientras que los ribosomas adheridos al RER están sintetizando proteínas que están destinadas a insertarse en una membrana, localizarse dentro de un orgánulo, o ser secretadas fuera de la célula por completo. El RER realiza modificaciones a las proteínas así como compartimentándolas. El SER cuenta la síntesis lipídica (por ejemplo, para hacer membranas) y las reacciones de desintoxicación entre sus funciones. Cabe señalar que los ribosomas en el RER no están permanentemente unidos, y después de haber producido una proteína, se disocian del RER y se reunen al acervo general de ribosomas libres en el citoplasma.
El complejo de Golgi, o cuerpos de Golgi, aunque físicamente independientes, son una extensión funcional al procesamiento y clasificación de proteínas que se produce en la sala de emergencias. Las proteínas salen del Golgi en vesículas unidas a la membrana celular u otros orgánulos. Las vesículas, aunque unidas a la membrana, generalmente no se cuentan como orgánulos: son simplemente pequeños paquetes de transporte.
Las mitocondrias son orgánulos complejos que no sólo están delimitados por una membrana, sino que también contienen una segunda membrana que está altamente almenada. Las mitocondrias hacen posible la respiración aeróbica, utilizando el oxígeno como oxidante para producir energía química (es decir, ATP) de manera mucho más eficiente que los procesos anaerobios utilizados por la mayoría de los procariotas. Esta capacidad de producir más energía a partir de la misma cantidad de alimento permite que las células eucariotas crezcan más grandes que las procariotas.
Los lisosomas son ácidos y contienen enzimas digestivas que descomponen las moléculas grandes de los alimentos —particularmente las proteínas y las grasas— para hacerlas utilizables por el resto de la célula. Estas enzimas funcionan de manera óptima en condiciones ácidas, lo que actúa como una especie de mecanismo de seguridad: si un lisosoma se rompe y libera sus enzimas en el citoplasma, no descompondrán los componentes celulares de ninguna manera porque el pH citoplásmico es cercano al neutro y las enzimas no funcionan bien. Una vez que se pensó que eran exclusivos de las células animales, ahora se han descrito lisosomas en todas las células de todos los reinos eucariotas.
Los peroxisomas también descomponen o convierten moléculas, pero generalmente actúan sobre moléculas más pequeñas por oxidación. Por ejemplo, algunos peroxisomas en las células hepáticas humanas se utilizan para descomponer el alcohol (etanol). Procesos como este suelen producir H 2 O 2, peróxido de hidrógeno, como subproducto. Dado que H 2 O 2 en altas concentraciones es dañino, los peroxisomas suelen contener una enzima, la catalasa, que la convierte en agua y oxígeno molecular.
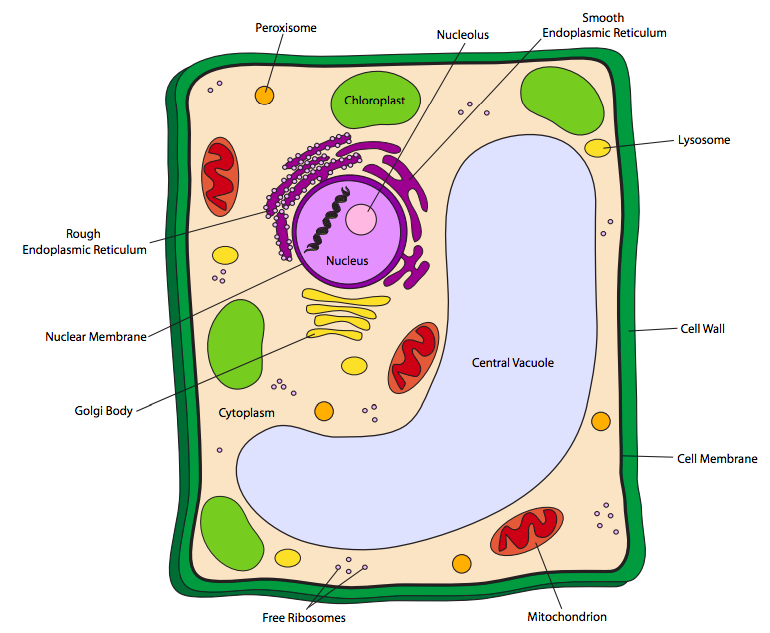
Las células vegetales tienen todos los orgánulos mencionados anteriormente, pero adicionalmente también pueden portar otros dos tipos de orgánulos: cloroplastos y vacuolas. Además de esto, las células vegetales también tienen una pared celular rígida externa a la membrana celular. Los cloroplastos son similares a las mitocondrias en forma y estructura (membranas dentro de la membrana externa). No obstante, su función es muy diferente: los cloroplastos absorben la energía luminosa del sol (u otras fuentes de luz) y las convierten en energía química en forma de azúcares simples para que la célula la almacene y use posteriormente, en esencia convirtiendo la célula en un almacén y centro de distribución de energía solar. Dado que las plantas y algunas bacterias fotosintéticas son los únicos organismos capaces de convertir la energía solar en una forma útil para las células vivas, son cruciales para la supervivencia de todas las demás vidas.
Las vacuolas son esencialmente unidades de almacenamiento. Pueden almacenar almidones para su uso como fuentes de energía cuando la luz solar no está disponible o cuando la fotosíntesis inmediata por sí sola no es suficiente para cubrir las necesidades energéticas de la célula. Otras vacuolas, como la representada anteriormente en la Figura\(\PageIndex{3}\), almacenan agua, lo que ayuda a que la celda mantenga la rigidez en combinación con la pared celular.
Las paredes celulares vegetales están compuestas de materiales muy diferentes a las paredes celulares bacterianas mencionadas anteriormente. Las paredes celulares vegetales están compuestas principalmente por el polímero de glucosa, celulosa, pero también contienen otros polisacáridos. Dependiendo del tipo de célula vegetal, puede haber múltiples capas de celulosa que componen la pared celular. La madera y la corteza de los árboles, por ejemplo, tienen tanto una pared celular primaria (delgada) como una pared secundaria (gruesa), mientras que las hojas solo tendrían una pared primaria. Los hongos también tienen paredes celulares, y ellos también son diferentes de las paredes celulares bacterianas. Los verdaderos hongos tienen paredes celulares que están compuestas principalmente por el polisacárido quitina, y ninguna celulosa.
Curiosamente, dos grupos una vez clasificados como hongos: oomicetos y dictyostelidos, tienen paredes celulares compuestas por celulosa (y algunos tienen ambas). Estos organismos han sido reclasificados en Protista.
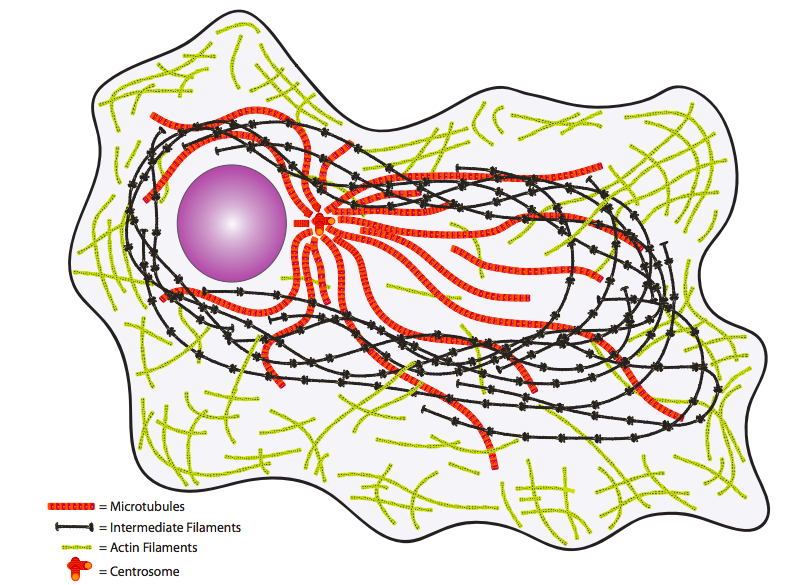
Por último, considere el citoplasma. Una vez considerado meramente el ambiente acuoso en el que flotaban las moléculas u orgánulos “importantes”, ahora se entiende mejor que está lleno de importantes elementos estructurales y de transporte (Figura\(\PageIndex{4}\)). El citoesqueleto proporciona no solo una estructura física interna sino también un sistema de transporte para mover moléculas, vesículas e incluso orgánulos a donde se necesitan.
Todas las partes celulares introducidas en este capítulo serán explicadas con mucho mayor detalle en capítulos posteriores. Más importante aún, las relaciones entrelazadas entre muchas de las moléculas y orgánulos serán discutidas y dilucidadas. Al pasar por este curso, notarás que las mismas especies aparecen una y otra vez como ejemplos. Estos son los organismos modelo en los que se basa la gran mayoría de la investigación en biología celular molecular. La mayoría de las investigaciones procariotas se han basado en Escherichia coli (E. coli), que es una bacteria Gram-negativa en forma de varilla que se encuentra comúnmente en el intestino de muchos animales superiores. La bacteria Gram-negativa del suelo, Bacillus subtilis, es un organismo formador de esporas que también se ha utilizado en la investigación porque, al igual que E. coli, su genoma es fácilmente manipulado para la experimentación, y también es relativamente fácil de cultivar en el laboratorio. En el lado eucariota, las levaduras (Saccharomyces cerevisiae, o Schizosaccharomyces pombe) son muy utilizadas para procesos intracelulares más simples debido a la genética simple y tiempos de generación muy rápidos. Caenorhabditis elegans (nematodo) y Drosophila melanogaster (mosca de la fruta) son organismos modelo invertebrados populares, especialmente para estudios genéticos y de desarrollo debido al pequeño número de células, en su mayoría con linaje trazable, y rápido tiempo de generación (para metazoos). Las ranas, particularmente la rana de garras sudafricanas, Xenopus laevis, y la rana leopardo del norte, Rana pipiens, son populares para ciertos tipos de estudios del desarrollo y del ciclo celular porque tienen ovocitos enormes que son susceptibles de muchos tipos de genética y fisiología manipulación no posible en otras celdas. Arabidopsis thaliana es el organismo modelo más utilizado para el estudio de la genética vegetal. Finalmente, debido a que son mamíferos como nosotros (los humanos), pero se reproducen rápidamente y pueden manipularse genéticamente con relativa facilidad, los ratones (Mus musculus) son muy utilizados en el estudio de mecanismos intra o intercelulares más complejos. Más recientemente, la secuenciación casi completa del genoma y el desarrollo de técnicas para manipularlo, han convertido a la rata (Rattus norvegicus) en otro organismo de investigación viable para el estudio de genes de mamíferos.
Puede encontrar más información sobre organismos modelo en el sitio web de los Institutos Nacionales de Salud de los Estados Unidos: www.nih.gov/science/models/
Los puntos en común que hacen que todos estos organismos sean excelentes modelos para el estudio de las moléculas de la célula y las interacciones entre ellas que constituyen la vida, son un tiempo de generación relativamente corto, genoma bien descrito (y en la mayoría de los casos completamente secuenciado) y facilidad de manipulación experimental. La mayoría de las moléculas y mecanismos que aprenderás en el curso fueron descubiertas en los mecanismos modelo más simples, y luego se volvieron a encontrar, muchas veces con elaboración en los más complejos.

Miniatura: Diagrama de una célula procariota típica. (Dominio público; LadyOfHats).