
5.3:5.3 El ciclo de TCA
- Page ID
- 53006

Entonces eres un joven y caliente eucariota que luce todo tipo de elegantes orgánulos membranosos internos, con la necesidad de demostrar tu valía mejor que los viejos procariotas guardianes — ¿qué haces? Bueno, ¡haz montones de ATP, claro! Y aparentemente sin esfuerzo en eso, usando solo las escoritas sobrantes después de que la glucólisis haya tomado su paso en una molécula de glucosa: NADH y piruvato. La glucólisis en eucariotas, como sus orígenes procariotas, ocurre en el citoplasma. El ciclo de TCA (también llamado ciclo del ácido cítrico) ocurre dentro de la matriz de las mitocondrias, un orgánulo de doble membrana.
El piruvato necesita abrirse camino desde el citoplasma, a través de las membranas mitocondriales externas e internas, y hacia la matriz mitocondrial. ¿Cómo funciona esto? La membrana externa es porosa, está plagada de grandes canales aniónicos relativamente inespecíficos conocidos como canales aniónicos dependientes de voltaje (VDAC), y admitirá fácilmente piruvato. Por el contrario, la membrana mitocondrial interna es altamente impermeable, y la entrada de piruvato está específicamente regulada por una proteína transportadora de piruvato.
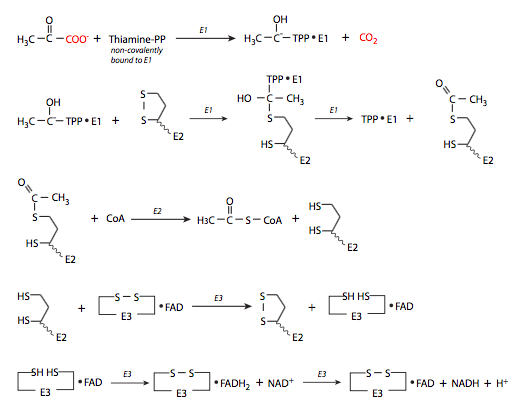
1. Una vez que el piruvato ingresa a la matriz mitocondrial, el complejo piruvato deshidrogenasa (compuesto por tres subunidades enzimáticas E1, E2 y E3) lo convierte en acetil-CoA (Figura\(\PageIndex{3}\)) para ingresar al ciclo del ácido tricarboxílico (TCA). Esta reacción genera NADH y libera CO 2.
El complejo piruvato deshidrogenasa (Figura\(\PageIndex{4}\)) es en realidad una amalgama de tres enzimas. Es decir, hay tres subunidades al complejo: piruvato deshidrogenasa (E1), dihidrolipoil transacetilasa (E2) y dihidrolipoil deshidrogenasa (E3). Estas tres subunidades están asociadas por enlaces no covalentes. La subunidad E1 de piruvato deshidrogenasa actúa primero, utilizando el cofactor pirofosfato de tiamina (TPP) para ayudar a eliminar un CO 2 del piruvato para generar hidroxietil-TPP.
Esto es utilizado inmediatamente como sustrato por E2, resultando en regeneración de TPP y reactivación de piruvato deshidrogenasa, y también haciendo el intermedio acetil-dihidrolipoamida. La coenzima A, que también es un sustrato para E2, tiene un grupo sulfhidrilo que ataca al grupo acetilo de la acetil-dihidrolipoamida. El grupo acetilo se transfiere inmediatamente a la Coenzima A para formar el acetil-CoA que entra en el ciclo de TCA.
El paso final es que la dihidrolipoamida sea oxidada de nuevo a lipoamida por E3. Es esta etapa de oxidación la que genera el NADH a partir de NAD +.
Ahora considere la descomposición de la glucosa. Recordemos que el desglose completo de ese azúcar de seis carbonos debería producir seis moléculas de dióxido de carbono único. En la glucólisis, la glucosa se descompone en dos moléculas de piruvato de tres carbonos. A medida que el piruvato se convierte en acetil-CoA, se genera un CO 2 por molécula de piruvato. Eso deja solo cuatro carbonos (en dos moléculas de 2 carbonos de acetil-CoA) fuera de la glucosa original 6. El ciclo de TCA liberará cada uno de esos carbonos como CO 2 también. Conocer las reacciones en las que se liberan los carbonos restantes es una buena manera de estudiar la primera mitad del ciclo de TCA.
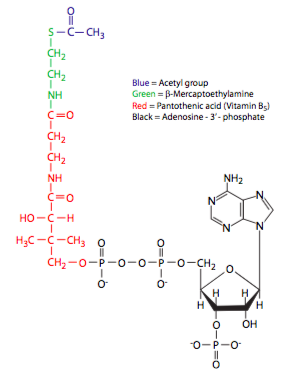
Como parte integral de la coenzima A, la vitamina B 5, o ácido pantoténico, es necesaria para el ciclo de TCA, y por lo tanto, para la generación eficiente normal de ATP. Sin embargo, a diferencia de otras vitaminas, la deficiencia de B 5 es rara, y generalmente se asocia con deficiencia en otras vitaminas o desnutrición general. Por otro lado, la deficiencia en otra vitamina B involucrada en la actividad piruvato deshidrogenasa (Figura\(\PageIndex{4}\)), la tiamina (B1), puede conducir a síntomas de enfermedad conocidos como beriberi.
El arsénico, o más específicamente los compuestos que contienen arsénico como el arsenito y el arseniato, son venenosos para las células al interferir con esta reacción. El compuesto de arsénico puede interactuar con la dihidrolipoamida, dando como resultado la ciclación mediante la unión de ambos sulfuros de sulfhidrilo al átomo de arsénico. Esto evita que E2 funcione, y no se puede generar acetil-CoA para la producción de ATP a través del ciclo de TCA y la fosforilación oxidativa. Cabe señalar que estos compuestos de arsénico también afectan a otros com- libras que contienen sulfhidrilo, y dentro del contexto del ciclo de TCA, también pueden inactivar α-cetoglutarato deshidrogenasa, que es similar a la piruvato deshidrogenasa.
Los síntomas del beriberi se clasifican en dos grupos: el beriberi húmedo afecta el sistema cardiovascular con síntomas como agrandamiento del corazón, congestión pulmonar, dificultad para respirar, hinchazón de la parte inferior de las piernas, insuficiencia cardíaca congestiva; beriberi seco (también conocido como síndrome de Wernicke-Korsakoff) afecta el sistema nervioso. Los síntomas incluyen polineuritis tanto en el sistema nervioso central como periférico, lo que lleva a dolor, hormigueo, pérdida de sensibilidad en las extremidades, pérdida de la función muscular o parálisis de la parte inferior de las piernas, vómitos, nistagmo, y eventualmente confusión mental, dificultades del habla, coma y muerte.
Las deficiencias genéticas en el complejo piruvato deshidrogenasa conducen a problemas similares, pero más inmediatamente graves. La mutación más común es una mutación dominante ligada a X en la subunidad a de E1. Las mutaciones de pérdida de función de PDC, así como las mutaciones en la piruvato carboxilasa y las mutaciones en la citocromo oxidasa, se consideran causas de la enfermedad de Leigh, que a menudo es mortal neonatalmente, aunque las excepciones han sobrevivido poco más de una década. La acidosis láctica grave y la incapacidad de generar suficiente energía, especialmente en las neuronas (que normalmente serían capaces de metabolizar la grasa -ver sección del catabolismo de ácidos grasos- pero no en estos pacientes) y las células musculares, es la causa subyacente de los síntomas.
2. El acetil-CoA ingresa al ciclo del ácido tricarboxílico como sustrato de la citrato sintasa, que lo agrega al oxaloacetato para producir citrato. Esta es la razón por la que a este ciclo también se le llama ciclo del ácido cítrico. El citrato, que tiene tres grupos carboxilo, es un ácido tricarboxílico, lo que lleva al nombre que utilizará este texto. El otro nombre común para esto es el ciclo de Krebs, ya que fue propuesto por primera vez por Hans Krebs en 1937.
La citrato sintasa es una enzima dimérica que en su forma nativa tiene una hendidura de unión para el oxaloacetato. La unión del oxaloacetato provoca un desplazamiento conformacional que cierra el sitio de unión al oxaloacetato, lo bloquea y revela simultáneamente el sitio de unión de acetil-CoA. El modelo actual para esta reacción implica tres etapas: acetil-CoA se convierte en un intermedio enol, que ataca al oxaloacetato para formar citronil-CoA (S-citril-CoA), que luego se hidroliza a citrato y coenzima A.
3. En el siguiente paso, la aconitasa reorganiza el citrato para hacer isocitrato.
La aconitasa empuja el citrato a un intermedio cis-aconitato, que luego se convierte en isocitrato. Curiosamente, si bien la aconitasa contiene un racimo de Fe-S, no parece participar en reacciones redox como suele ser el caso de dichos grupos. En cambio, su propósito es mantener el cis-aconitato en su lugar dentro de la enzima, ya que [el cis-aconitato] sufre un extraño giro molecular en su camino hacia el isocitrato.
El fluoroacetato de sodio, también conocido como compuesto 1080, es un pesticida común que se usa principalmente contra plagas de roedores y otros mamíferos, y puede actuar en humanos si se ingiere. Una vez introducido en el organismo, se puede convertir en fluoroacetil-CoA y luego en fluorocitrato, que luego actúa como inhibidor competitivo de la aconitasa. Como tal, la intoxicación afecta de manera más grave y rápida a los tejidos con altas necesidades energéticas. No se reconocen antídotos efectivos para las dosis letales de intoxicación por fluoroacetato.
4. El isocitrato es un sustrato para la isocitrato deshidrogenasa, que transfiere un electrón de alta energía del isocitrato a NAD + para hacer NADH y α-cetoglutarato. Esta reacción también libera un CO 2. Para aquellos que hacen un seguimiento en casa, eso deja dos carbonos más de los seis en glucosa.
La isocitrato deshidrogenasa dependiente de NAD + se encuentra en dos isoformas en mamíferos: una isoforma que utiliza NAD + en la matriz mitocondrial y una isoforma que usa NADP + que se encuentra en el citosol y en las mitocondrias. La reacción comienza con la oxidación generadora de NADH de isocitrato a oxoalosuccinato, que luego se descarboxila con la ayuda de un cofactor Mn 2+ o Mg 2+ para liberar el dióxido de carbono y formar α-cetoglutarato.
5. El alfa-cetoglutarato también se oxida (por α-cetoglutarato deshidrogenasa) generando NADH y succinil-CoA. Al igual que acetil-CoA, este compuesto asociado a CoA contiene un enlace tioéster de alta energía. Esta reacción libera el CO 2 final de la glucosa.
La α-cetoglutarato deshidrogenasa es muy similar al complejo piruvato deshidrogenasa estructural y mecanísticamente. Existen tres enzimas: la α-cetoglutarato deshidrogenasa, una dihidrolipoil transsuccinilasa y dihidrolipoil deshidrogenasa. También similar al complejo piruvato deshidrogenasa, el producto final es una molécula que contiene un enlace tioéster de alta energía.
6. El CoA es regenerado por succinil-CoA sintetasa, que también forma succinato y GTP o ATP. Este GTP es energéticamente un ATP equivalente, y hecho en células animales. Los homólogos bacterianos y vegetales de esta enzima usan ADP y producen ATP. La formación de este ATP/GTP es posible porque se rompe el enlace tioéster de alta energía de succinil-CoA.
La succinil-CoA sintetasa primero reúne la succinil-CoA y el fosfato inorgánico (en solución dentro de la matriz mitocondrial así como el citosol) para producir fosfato de succinilo y liberar el CoA. Después el fosfato se transfiere del fosfato de succinilo, a la propia enzima temporalmente, que luego deja caer el succinato. Y finalmente, el fosfato se transfiere a GDP/ADP.
7. A continuación, se oxida el succinato. La enzima que hace esto, la succinato deshidrogenasa, es un poco diferente de las otras deshidrogenasas porque esta pasa a estar incrustada en la membrana mitocondrial interna, y en lugar de transferir el electrón a NAD +, el electrón se transfiere a FAD, haciendo FADH 2, y fumarato. La energía en FADH 2 también se puede utilizar para alimentar la producción de ATP similar a la energía en NADH.
A pesar de que la simplificación habitual de clase intro-clase es que FADH 2 es aproximadamente un equivalente al NADH, la situación es en realidad más complicada. A diferencia del NAD +, y para el caso, a diferencia de la mayoría de las incidencias de FAD, el FAD se une covalentemente a la succinato deshidrogenasa. Por lo tanto, no es un metabolito soluble, ni está disponible para ser reoxidado como el NADH. Está, por supuesto, reoxidado. Pero, esto ocurre dentro del contexto de la cadena de transporte de electrones (donde se le conoce como Complejo II), con la ayuda de la Coenzima Q.
8. La fumarasa cataliza la adición de agua al fumarato para generar malato.
El doble enlace carbonado del fumarato es atacado por un hidroxilo (OH -) para formar una transición carbaniónica, que luego toma un protón (H +) de la enzima para formar el malato. La enzima fumarasa es protonada al mismo tiempo que se une al fumarato, y es desprotonada al final para formar el malato.
9. El malato es oxidado por malato deshidrogenasa en la reacción final del ciclo de TCA para generar más NADH, y oxaloacetato, el último de los cuales se puede agregar a acetil-CoA para iniciar el ciclo de nuevo.
La malato deshidrogenasa es similar en estructura a la lactato deshidrogenasa y la alcohol deshidrogenasa mencionadas en la sección de fermentación. Energéticamente, el cambio estándar de energía libre de esta reacción es muy positivo (29.7 kJ/mol) pero el oxaloacetato se convierte rápidamente en citrato, por lo que se favorece una mayor formación de oxaloactetato sobre la formación de malato.
Ahora que se ha descrito el ciclo completo, cabe señalar que la regulación de este ciclo es principalmente a través de la disponibilidad de acetil-CoA y oxaloacetato, así como la concentración de NADH. A medida que aumenta la tasa de respiración, los niveles de NADH bajan a medida que se oxidan para producir ATP (ver siguiente sección). Esta caída en [NADH] provoca entonces un aumento en el oxaloactetato, que luego es utilizado por la citrato sintasa. [Acetil-CoA] está regulado por su síntesis por piruvato deshidrogenasa. En el reverso de la regulación, tanto el NADH como la succinil-CoA son fuertes inhibidores de la α-cetoglutarato deshidrogenasa. Así, a medida que se agota el NADH, la enzima se desinhibe y aumenta su producción de más NADH.
Habiendo tomado los 2 piruvatos creados durante la glucólisis a través del ciclo de TCA para completar la oxidación en CO 2, ¿qué le queda a nuestro intrépido héroe eucariota? Dos equivalentes de ATP (GTP) seis NADH y dos FADH 2. Esto apenas parece ser un tesoro de energía utilizable que valga la pena presumir. Afortunadamente, la mitocondria no está terminada. A continuación, los electrones de alta energía darán un paseo en la cadena de transporte de electrones, y a través de la magia de la fosforilación oxidativa, producirán ATP por el cubo.