
6.2: El Ciclo Calvino
- Page ID
- 53154

Las reacciones oscuras (fijación de carbono) varían según el tipo de planta. El conjunto más común de reacciones de fijación de carbono se encuentra en plantas de tipo C3, las cuales se denominan así porque el principal intermedio estable es la molécula de 3 carbonos, gliceraldehído-3-fosfato. Estas reacciones, mejor conocidas como el ciclo de Calvin (Figura\(\PageIndex{6}\)), fijan CO 2 sobre la pentosa, ribulosa 1,5-bis-fosfato (RuBP).
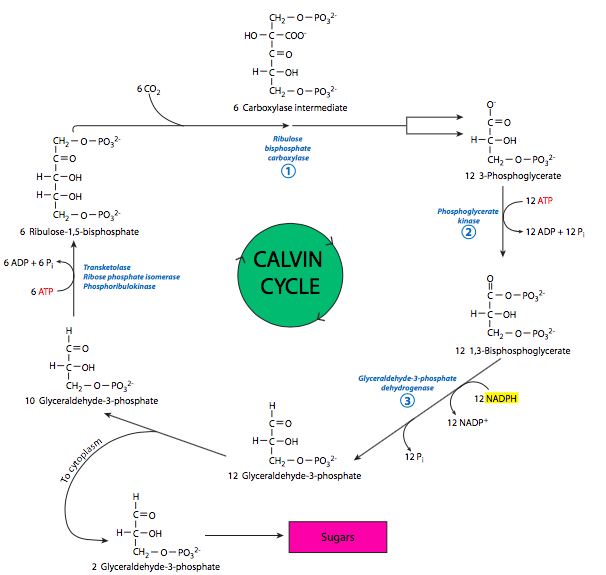
La parte de producción del ciclo comienza con la formación de RuBP a partir de gliceraldehído-3-fosfato. Luego, se produce el paso de limitación de velocidad:
Ribulosa 1,5-bisfosfato y CO2 se unen por rubisco. Las carboxilasas son enzimas relativamente lentas como familia, y rubisco es una de las más lentas. Se forma un intermedio de 6 carbonos pero es inestable, y se descompone rápidamente para producir dos moléculas de 3-fosfoglicerato. Algunas enzimas familiares (de la glucólisis, aunque esto está sucediendo en el estroma, no en el citoplasma) ahora entran en juego.
La fosfoglicerato quinasa fosforila 3-PG a 1,3-bisfosfosglicerato. El 1,3-BPG se reduce luego por gliceraldehído-3-fosfato deshidrogenasa para formar gliceraldehído-3-P. Este paso requiere la energía liberada por la oxidación del NADPH. Una pequeña porción (1/6 º) del GAP que se elabora se exporta luego del cloroplasto y se utilizará para formar carbohidratos más complejos. Sin embargo, la mayoría se recicla a través de la fase de recuperación del ciclo de Calvin para regenerar el NADP.
Como si tener una enzima central que se mueve al ritmo de un caracol y necesitar reciclar la mayor parte de su producto potencial no fuera lo suficientemente malo, las plantas C3 también tienen que enfrentarse al secuestro de rubisco para un conjunto de reacciones competidoras y desperdiciadoras de energía conocidas como fotorespiración. Bajo condiciones de CO 2 bajo y alto O 2 en la atmósfera local, el oxígeno, en lugar de dióxido de carbono, se une al rubisco y forma 3-PG y 2-fosfoglicolato a partir de su reacción con RubP. Como se detalla en la Figura\(\PageIndex{7}\), el 2-fosfoglicolato se desfosforila a glicolato y se transporta fuera del cloroplasto. A partir de ahí, sufre una serie de reacciones en los peroxisomas y mitocondrias para transformarlo en 3-PG, que luego puede ir en el cloroplasto y participar en el ciclo Calvino. Desafortunadamente para la célula, en el transcurso de estas reacciones, se utilizan NADH y ATP, disminuyendo así la disponibilidad de energía dentro de la célula.
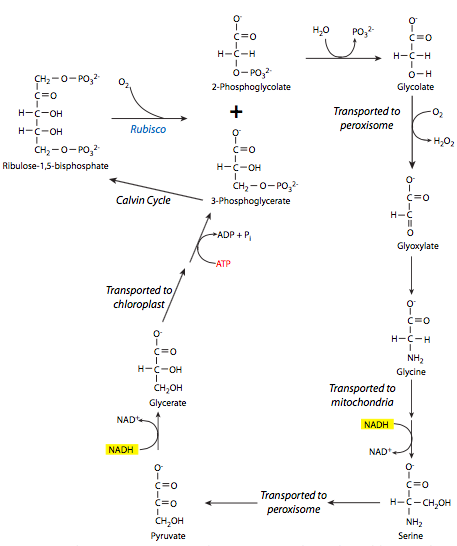
Esto es un problema particular en climas cálidos, debido a que la actividad oxigenasa del rubisco aumenta más que la actividad de carboxilasa a medida que aumenta la temperatura. Esto lleva a un efecto secundario interesante: en las plantas C3, a medida que aumenta la temperatura y el CO 2 es superado por O 2 para la unión de rubisco, los estomas de las hojas necesitan permanecer abiertos por más tiempo para permitir la adquisición de suficiente CO 2 de la atmósfera. Esto a su vez permite que se escape más vapor de agua del interior de la celda, lo que lleva a la deshidratación. Por lo tanto, las plantas C3 se encuentran en desventaja competitiva en climas cálidos y secos en comparación con las plantas que no utilizan rubisco para la fijación de carbono.
¿Qué pasa con las plantas adaptadas a tales climas? Las plantas C4, que incluyen algunas gramíneas, maíz, caña de azúcar y malezas, utilizan PEP carboxilasa (que no tiene las molestas capacidades fotorrespiratorias del rubisco y una mayor afinidad por el CO 2) para fijar el dióxido de carbono a PEP, haciendo oxaloacetato. En un giro interesante, el oxaloactetato, después de la conversión a malato, es descarboxilado para dar de nuevo CO 2, que se alimenta al rubisco y al ciclo de Calvino.
La PEP carboxilasa realmente fija HCO3 - a PEP en lugar de CO 2 directamente. El CO 2 atmosférico se convierte en bicarbonato por la anhidrasa carbónica.
El mecanismo C4, también llamado vía Hatch-Slack, utiliza dos conjuntos de células, una capa externa (mesófila) que toma aire y fija el CO 2 a PEP y produce malato, y una capa interna de células (vaina de haz) que toma el malato, y lo descarboxila para su enzima rubisco. Las dos células están conectadas a través de plasmodesmas (ver capítulo Interacciones célula-célula). Aunque energéticamente más cara que la fijación de carbono por plantas C3 en climas más fríos, la vía C4 supera a C3 en eficiencia a medida que aumentan las temperaturas y aumenta la fotorespiración.
Se han encontrado variaciones de esta vía en las que el aspartato se transporta a las células de la vaina del haz en lugar de malato. Después de la descarboxilación del malato por la enzima málica (dependiente de NAD en algunas especies, dependiente de NADP en otras) para liberar el CO 2 para rubisco, el piruvato resultante se transporta de regreso a la célula del mesófilo donde es fosforilado por la piruvato-fosfato diquinasa para generar PEP para reentrada en el Ciclo C4.
Las plantas del desierto van un paso más allá que las plantas C4. Al vivir en ambientes extremadamente calurosos y secos durante el día, pero relativamente fríos por la noche, muchas suculentas del desierto (como los cactus) son diurnas, y solo abren sus estomas por la noche (cuando las temperaturas son significativamente más bajas y el agua se evapora mucho más lentamente) para la recolección de CO 2, que es entonces fijado a través de la vía CAM al malato. Después en las horas de luz, el CO 2 se libera del malato y se usa en el ciclo Calvino para generar carbohidratos.
La vía del metabolismo del ácido crassuláceo (CAM)
La vía del metabolismo del ácido crassuláceo (CAM) se llama así por una vía de fijación de carbono descubierta en la familia Crassulaceae de plantas suculentas incluyendo piñas, así como varias especies de cactus. Utiliza un mecanismo bioquímico similar al de la ruta C4, pero ocurre dentro de una sola célula fotosintética. La mayor diferencia es que el CO 2 solo se toma por la noche, y rápidamente se convierte en malato, el cual se almacena en vacuolas hasta el día. El malato es entonces liberado y descarboxilado para proporcionar a la RubP carboxilasa (rubisco) una corriente constante de CO 2 para su fijación. Debido a que existe tal avalancha de actividad de PEP carboxilasa por la noche para fijar el CO 2 atmosférico a PEP, existe una alta tasa de descomposición del almidón para proporcionar la glucosa para la generación glicolítica de PEP. Curiosamente, como el malato es descarboxilado en el día, su producto, piruvato, puede ser utilizado para resintetizar glucosa (ver sección de gluconeogénesis) y luego almidón.