
6.5: Síntesis de glucógeno
- Page ID
- 53189

Aunque la glucosa es el combustible principal para las células, no es una molécula eficiente para el almacenamiento a largo plazo en organismos complejos (es decir, mayores que unicelulares). Por lo tanto, tanto en plantas como en animales, las moléculas de glucosa se unen entre sí para formar polisacáridos conocidos como glucanos. En los animales, el glucano formado es glucógeno, el cual consiste en moléculas de glucosa unidas por enlaces glucosídicos α (1->4), y enlaces ramificados α (1->6) aproximadamente entre 8 y 14 residuos separados. El tamaño promedio de una unidad de glucógeno es un gránulo citoplásmico que contiene más de 100000 moléculas de glucosa. La adición de una glucosa-1-fosfato a otra (o a una cadena de glucógeno) es energéticamente desfavorable, por lo que debe acoplarse con una reacción suficientemente exergónica para proceder.
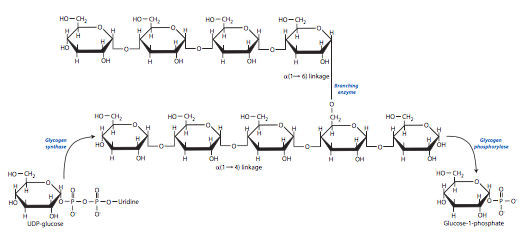
La síntesis de glucógeno comienza con UDP-glucosa fosforilasa, que combina el nucleótido uridina trifosfato (UTP) con glucosa-1-fosfato para liberar pirofosfato (PP i) y formar UDP-glucosa.
La reacción de intercambio de fosfoanhídrido catalizada por UDP-glucosa fosforilasa es mínimamente exergónica. Sin embargo, el pirofosfato liberado es hidrolizado rápidamente por la pirofosfatasa inorgánica, una enzima citosólica ubicua, en una reacción altamente exergónica. Esta hidrólisis de pirofosfato es un mecanismo utilizado en muchas vías biosintéticas para proporcionar energía para reacciones de otro modo endergónicas.
En el siguiente paso, la glucógeno sintasa une la UDP-glucosa a la cadena de glucógeno preexistente con un enlace α (1->4). No puede unir dos glucosas individuales, solo se suman a una cadena preexistente. Esto significa que debe haber alguna solución alternativa para las dos primeras glucosas:la glucogenina es una enzima que cataliza la adición de UDP-glucosa a sí misma, y puede hacerlo hasta por siete moléculas de UDP-glucosa, formando así un cebador corto para que la glucógeno sintasa funcione. Además, la glucógeno sintasa solo puede agregar glucosas con un enlace α (1->4). Para que ocurra la ramificación, se necesita una enzima de ramificación (específicamente, amylo- (1,4->1,6) -transglicosilasa. Esta enzima puede transferir segmentos de cadena terminal al hidroxilo de 6 carbonos de cualquier glucosa en una cadena de glucógeno. Sin embargo, las ramas solo se pueden agregar si hay al menos 4 residuos de glucosa entre ellas, y si la cadena originaria tenía al menos 11 residuos de longitud.
Síntesis de oligosacáridos
Al igual que la síntesis de glucógeno, la síntesis de oligosacáridos también requiere el paso inicial de acoplar el azúcar con un nucleótido. En los mamíferos, un disacárido principal es la lactosa, que es el enlace de una galactosa y una glucosa, y la formación es catalizada por la lactosa sintasa. Sin embargo, antes de que la lactosa sintasa sea capaz de actuar, la galactosa debe estar primero en forma de UDP-galactosa. De igual manera, en las plantas, el disacárido principal es la sacarosa, formada por la unión de UDP-glucosa y fructosa-6-fosfato. Esto da como resultado sacarosa-6-fosfato, que luego se desfosforila fácilmente a sacarosa. Este tipo de mecanismos también se utilizan en la glicosilación de proteínas y lípidos, los cuales serán discutidos principalmente en el capítulo de procesamiento y tráfico de proteínas.
La mutación de galactosa-1-fosfato uridiltransferasa o mutaciones de otras enzimas en esta vía (las mutaciones de uridiltransferasa son las más comunes y generalmente las más graves) pueden conducir a galactosemia, una enfermedad genética humana cuyos síntomas comienzan en la infancia y pueden incluir retraso mental, daño hepático, ictericia, vómitos y letargo. La causa de estos síntomas es generalmente una acumulación de galactosa-1-fosfato, especialmente en el hígado y el tejido nervioso. Afortunadamente, con un diagnóstico precoz, los síntomas se pueden prevenir evitando los productos lácteos (lactosa).
Las principales especies de hexosa además de la glucosa son fructosa, manosa y galactosa. La interconversión entre estas hexosas puede ocurrir a través de intermedios, como se demuestra en la glucólisis (glucosa-6-P a fructosa-6-P). La manosa-6-P se puede convertir en fructosa-6-P mediante fosfomanosa isomerasa. La galactosa se puede convertir de manera similar, en galactosa-1-P y luego en glucosa-1-P. La conversión de galactosa a glucosa también puede realizarse por epimerización de UDP-glucosa a UDP-galactosa vía redox intermedio usando NAD +/NADH.