
7.3: Replicación procariota
- Page ID
- 53013

La replicación del ADN comienza en un origen de replicación. Solo hay un origen en los procariotas (en E. coli, ORic) y se caracteriza por matrices de secuencias repetidas. Estas secuencias se envuelven alrededor de una proteína de unión al ADN, y al hacerlo, ejercen presión sobre los enlaces H entre las hebras del ADN, y el cromosoma comienza a descomprimirse en un área rica en AT envuelta alrededor de esta proteína. Recuerde que los pares A-T son 33% más débiles que los pares G-C debido a la menor cantidad de enlaces de hidrógeno. El uso de extensiones de ADN ricas en AT como puntos de separación de cadenas es un tema recurrente a través de una variedad de operaciones de ADN. La separación de las dos cadenas es bidireccional, y las ADN polimerasas actuarán en ambas direcciones para terminar el proceso lo más rápido posible. La velocidad es importante aquí porque mientras se produce la replicación, el ADN es vulnerable a la rotura, y la mayoría de los procesos metabólicos se cierran para dedicar la energía a la replicación. Incluso en los procariotas, donde las moléculas de ADN son órdenes de magnitud más pequeñas que en los eucariotas, el tamaño de la molécula de ADN cuando se desentraña de las proteínas de empaquetamiento protectoras la hace altamente susceptible al daño físico solo por los movimientos de la célula.
La primera proteína de unión a ORic, ADNA, se une a cajas de ADN, las cuales son segmentos de 9 pares de bases con una secuencia consenso de TTATCCACA. ORic tiene cinco de estas repeticiones, y una proteína de ADN se une a cada una de ellas. HU e IHF son proteínas similares a histonas que se asocian con el ADN y juntas doblan esa parte del ADN en un bucle circular, situándolo justo sobre la otra característica principal de ORic, las repeticiones ricas en AT de 13 pb (GATCTNTNTNTTTTTT). El ADN hidroliza ATP y rompe los enlaces H entre las cadenas en las repeticiones 13mer, también conocido como fusión del ADN. Esto permite complejos de DnaB [y DnaC, que es una proteína de carga que ayuda a unir DnaB (6) a la cadena con la hidrólisis acompañante de ATP. Además, se reclutan cinco ADN más para estabilizar el bucle.] para unirse a cada región monocatenaria del ADN en lados opuestos de la burbuja de replicación recién abierta.
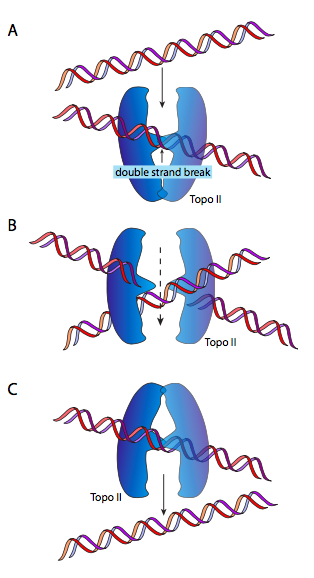
DnaB es una helicasa; su actividad enzimática es descomprimir/desenrollar el ADN por delante de la ADN polimerasa, para darle ADN monocatenario para leer y copiar. Lo hace en asociación con proteínas de unión a ADN monocatenario (SSB) y ADN girasa. La función de SSB es casi autoexplicativa: el ADN monocatenario es como el ARN en su capacidad para formar estructuras secundarias complejas por emparejamiento interno de bases, por lo que SSB evita eso. La ADN girasa es una topoisomerasa tipo II, y tiene la tarea de introducir superenrollamiento negativo en el ADN. Esto es necesario porque la descompresión del ADN por helicasa también lo desenrolla (ya que es una doble hélice) y provoca la introducción de superenrollamiento positivo. Esto significa que toda la molécula circular se retuerce sobre sí misma: imagínese sosteniendo una banda elástica en dos manos y retorciéndola. A medida que se acumula el superenrollamiento, el ADN se enrolla más fuertemente, hasta el punto de que sería imposible que la helicasa lo descomprima. DNAB/girasa puede aliviar este estrés cortando temporalmente el ADN bicatenario, pasando un bucle de la molécula a través de la brecha y resellándolo.
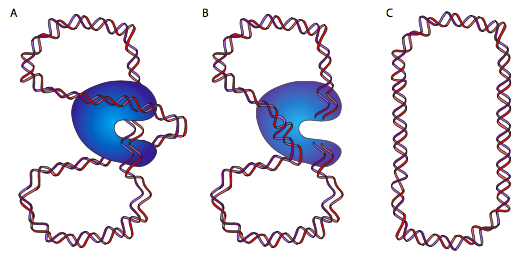
Esto (ojalá) tenga mucho más sentido mirando el diagrama. O, volviendo a nuestra goma elástica, dale a la banda de goma un giro o dos, luego pega los dos extremos con cinta adhesiva. Si corta la banda de goma y pasa una porción adyacente de la banda elástica a través de ese recorte, luego vuelve a conectar los extremos cortados, encontrará que hay un giro menos. Ingenioso, ¿eh? En este punto, algunos de ustedes van a decir, pero si tuercen una banda de goma flotante libre, como uno podría imaginar un cromosoma de ADN circular flotante en E. coli, esperaría que se desenroscara naturalmente. Técnicamente, sí, pero debido a la gran masa del cromosoma, su asociación con diversas proteínas y la membrana celular, y la viscosidad de su entorno, no se comporta como si fuera completamente libre.
Una vez que se ha abierto ORic y las helicasas se han unido a los dos lados de la horquilla de replicación, la máquina de replicación, también conocida como el replisoma, puede comenzar a formarse. Sin embargo, antes de que las ADN polimerasas tomen posiciones, necesitan ser cebadas. Las ADN polimerasas son incapaces de unir dos nucleótidos libres individuales para comenzar a formar un ácido nucleico; solo pueden agregarse a una cadena preexistente de al menos dos nucleótidos. Por lo tanto, una ARN polimerasa especializada (RNAP's no tienen esta limitación) conocida como primasa es una parte del replisoma, y las lecturas crean una cadena corta de ARN denominada el cebador para que la ADN polimerasa se agregue. Aunque solo se necesitan unos pocos nucleótidos, los cebadores procariotas pueden tener hasta 60 nt dependiendo de la especie.
Hasta la fecha se han descubierto al menos cinco ADN polimerasas procariotas. La ADN polimerasa primaria para replicación en E. coli es la ADN Polimerasa III (Pol III). Pol I también está involucrado en el mecanismo básico de replicación del ADN, principalmente a ll en brechas creadas durante la síntesis de cadenas rezagadas (definidas 3 páginas más adelante) o a través de mecanismos de corrección de errores. La ADN polimerasa II y los recientemente descubiertos Pol IV y Pol V no participan en la replicación cromosómica, sino que se utilizan para sintetizar ADN cuando se necesitan ciertos tipos de reparación en otros momentos del ciclo de vida celular.
La ADN polimerasa III es una holoenzima multisubunidad, con subunidades α, ε y θ que comprenden la polimerasa central, y τ, γ, δ, δ ', χ, ψ y β que se unen para formar la holoenzima completa. La polimerasa central tiene dos actividades: la subunidad α es la función de la polimerasa, leyendo una cadena de ADN y sintetizando una cadena complementaria con gran velocidad, alrededor de 150 nt/seg; la subunidad ε es una exonucleasa 3'-5' de “corrección” y actúa como corrector inmediato, eliminando la último nucleótido si es incorrecto. Esta prueba- lectura no llega más atrás: solo actúa sobre el nucleótido agregado más recientemente para corregir la incorporación errónea. Se utilizan otros mecanismos y enzimas para corregir las lesiones de ADN que surgen en otros momentos. [Como cuestión de nomenclatura, las exo nucleasas solo cortan los nucleótidos del ADN o ARN de cualquiera de los extremos, pero no en el medio. Las endo nucleasas escinden enlaces fosfodiéster localizados más profundamente dentro de una cadena de ácido nucleico.] La subunidad θ no tiene actividad enzimática y regula la función exonucleasa. A pesar de que tiene actividad polimerasa, la polimerasa central de Pol III tiene poca procesividad, es decir, solo puede sumar hasta 15 nucleótidos antes de disociarse del ADN molde. Dado que los genomas de cepas de E. coli tienen un promedio de cerca de 5 millones de pares de bases, la replicación en pequeños segmentos de 15 nt sería extraordinariamente ineficiente.
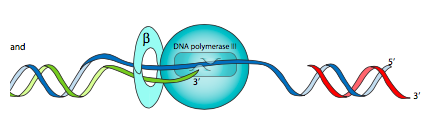
El complejo de cargador de pinza es un conjunto de ATPasa que se une a la unidad β-clamp tras la unión de ATP (pero la actividad de ATPasa no está activada). Cuando el complejo luego se une al ADN, activa la ATPasa, y la hidrólisis resultante de ATP conduce a cambios conformacionales que abren la pinza temporalmente (para rodear o alejarse de la cadena de ADN), y luego disociar el cargador de abrazadera del conjunto de abrazadera.
Aquí es donde se necesita la subunidad β. También conocida como la abrazadera β, es un dímero de subunidades semicirculares que tiene un orificio central a través del cual se enhebra el ADN. La polimerasa central, a través de una interacción α-β, se une a esta pinza β para que permanezca en el ADN por más tiempo, aumentando la procesividad de Pol III a más de 5000 nt. La pinza β se carga en (y se descarga de) el ADN mediante un complejo cargador de pinza (también llamado complejo γ) que consiste en subunidades γ (x3), δ, δ', χ y ψ.
La burbuja de replicación tiene dos horquillas de replicación: una vez que el ADN se abre (descomprime) en el origen, se puede formar una máquina de replicación en cada extremo, con las helicasas dirigiéndose en direcciones opuestas. Para la simplificación, consideraremos solo una bifurcación —que se abre de izquierda a derecha— en esta discusión con el entendimiento de que lo mismo está sucediendo con la otra bifurcación, pero en sentido contrario.
Lo primero que hay que notar al mirar un diagrama de una horquilla de replicación (Figura\(\PageIndex{11}\)) es que las dos porciones monocatenarias del ADN molde son antiparalelas. Esto no debería sorprender en este punto del curso, pero sí introduce un problema mecánico interesante. La helicasa abre el ADN bicatenario y conduce el resto de la máquina de replicación. Entonces, en la región monocatenaria al final de la helicasa, si miramos de izquierda a derecha, una cadena molde es de 3' a 5' (en azul), mientras que la otra es de 5' a 3' (en rojo). Dado que sabemos que los ácidos nucleicos se polimerizan añadiendo el fosfato 5' de un nuevo nucleótido al hidroxilo 3' del nucleótido anterior (5' a 3', en verde), esto significa que una de las cadenas, llamada cadena líder, se está sintetizando en la misma dirección que la máquina de replicación se mueve. No hay problema ahí.
La otra cadena es problemática: mirada linealmente, la hebra recién sintetizada iría de 3' a 5' de izquierda a derecha, pero las ADN polimerasas no pueden agregar nucleótidos de esa manera. ¿Cómo resuelven las células este problema? Se han propuesto varias posibilidades, pero aquí se representa el modelo actual. La máquina de replicación consiste en la helicasa, primasas y dos holoenzimas de ADN polimerasa III que se mueven en la misma dirección física (siguiendo a la helicasa). De hecho, los complejos pol III están físicamente unidos a través de subunidades τ.
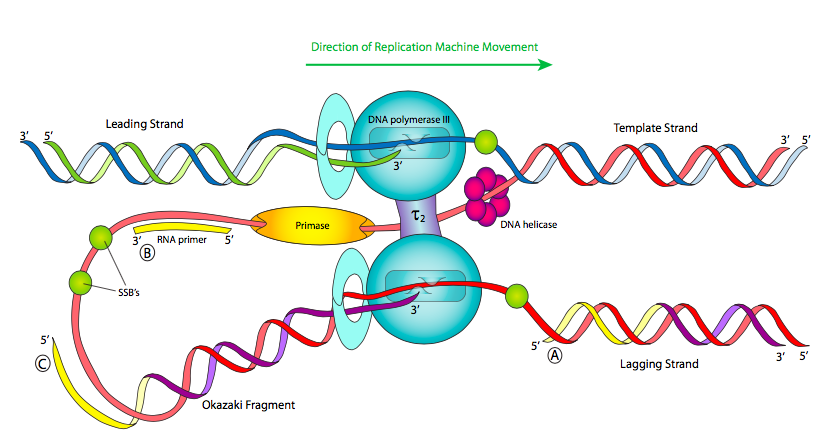
Para que la cadena molde que está 5' a 3' de izquierda a derecha se replique, la cadena debe alimentarse hacia atrás en la polimerasa. Esto se puede lograr ya sea dando la vuelta a la polimerasa o enrollando el ADN alrededor. Como muestra la Figura, el modelo actual es que la primasa también se mueve de izquierda a derecha, por lo que tiene poco tiempo para sintetizar rápidamente un cebador corto antes de tener que avanzar con el replisoma y comenzar de nuevo, dejando cebadores intermitentes a su paso. Debido a esto, Pol III se ve obligado a sintetizar sólo fragmentos cortos del cromosoma a la vez, llamados fragmentos de Okazaki después de su descubridor. Pol III comienza a sintetizar añadiendo nucleótidos en el extremo 3' de un cebador y continúa hasta que alcanza el extremo 5' del siguiente cebador. No conecta (y no puede) la hebra que está sintetizando con el extremo 5' del cebador.
La replicación del ADN se denomina proceso semidiscontinuo porque mientras la cadena principal se sintetiza continuamente, la hebra rezagada se sintetiza en fragmentos. Esto lleva a dos problemas principales: primero, quedan pequeños fragmentos de ARN en las cadenas recién hechas (justo en el extremo 5' para la cadena principal, en muchos lugares para la rezagada); y segundo, Pol III solo puede agregar nucleótidos libres a un fragmento de ADN monocatenario; no puede conectar otro fragmento. Por lo tanto, la nueva “hebra” no es entera, sino plagada de faltantes enlaces fosfodiéster.
El primer problema lo resuelve la ADN polimerasa I. A diferencia de Pol III, Pol I es una proteína monomérica y actúa sola, sin proteínas adicionales. También hay 10-20 veces más moléculas de Pol I que moléculas de Pol III, ya que son necesarias para tantos fragmentos de Okazaki. La ADN Polimerasa I tiene tres actividades: (1) como Pol III, puede sintetizar una cadena de ADN basada en un molde de ADN, (2) también como Pol III, es una exonucleasa correctora de 3'-5', pero a diferencia de Pol III, (3) también es una exonucleasa 5'-3'. La actividad exonucleasa 5'-3' es crucial en la eliminación del cebador de ARN (Figura\(\PageIndex{12}\)). La exonucleasa 5'-3' se une al ADN bicatenario que tiene una ruptura monocatenaria en la cadena principal del fosfodiéster como lo que sucede después de que los fragmentos de Okazaki se hayan sintetizado de un cebador al siguiente, pero no se puede conectar. Esta exonucleasa 5'-3' elimina entonces el cebador de ARN. La actividad polimerasa luego agrega nuevos nucleótidos de ADN al fragmento de Okazaki aguas arriba, llenando el hueco creado por la eliminación del cebador de ARN. La exonucleasa correctora actúa igual que lo hace para Pol III, eliminando inmediatamente un nucleótido incorrecto recién incorporado. Después de la corrección, la tasa de error general de incorporación de nucleótidos es aproximadamente de 1 en 107.
Técnicamente, la exonucleasa 5'-3' escinde el ADN en una región bicatenaria aguas abajo de la mella, y luego puede eliminar cualquier lugar de 1-10nt a la vez. Experimentalmente, la actividad exonucleasa 5'-3' puede ser escindida del resto de Pol I por la proteasa tripsina. Esto genera el “fragmento Klenow” que contiene la polimerasa y la exonucleasa correctora 3'-5'.
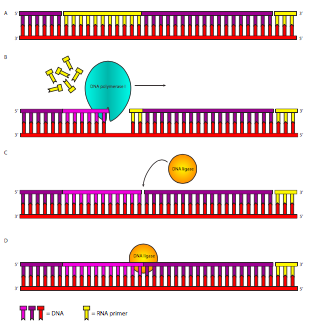
Figura\(\PageIndex{12}\). Síntesis de hebras rezagadas. Después de que la ADN polimerasa III haya extendido los cebadores (amarillo), la ADN polimerasa I elimina el cebador y lo reemplaza añadiéndolo al fragmento anterior. Cuando termina de eliminar el ARN y reemplazarlo con ADN, deja el ADN con un enlace fosfodiéster faltante entre el ADN sintetizado de pol III aguas abajo y el ADN sintetizado de pol I aguas arriba. Esta ruptura en la cadena principal de azúcar-fosfato es reparada por la ADN ligasa.
A pesar de que el ARN ha sido reemplazado por ADN, esto todavía deja una hebra fragmentada. Finalmente aparece el último jugador importante en la historia de la replicación del ADN: la ADN ligasa. Esta enzima tiene una tarea sencilla pero crucial: cataliza el ataque del 3'-OH a partir de un fragmento sobre el fosfato 5' del siguiente fragmento, generando un enlace fosfodiéster. Esta reacción requiere energía en forma de hidrólisis de ATP o NAD + dependiendo de la especie (E. coli usa NAD +) generando AMP y PP i o NMN +.