11.3: Plegamiento de proteínas en el retículo endoplásmico
- Page ID
- 53102

La luz del retículo endoplásmico (ER) juega cuatro papeles principales en el procesamiento de proteínas:
- plegado/replegamiento del polipéptido,
- glicosilación de la proteína,
- ensamblaje de proteínas de múltiples subunidades, y
- empaquetamiento de proteínas en vesículas.
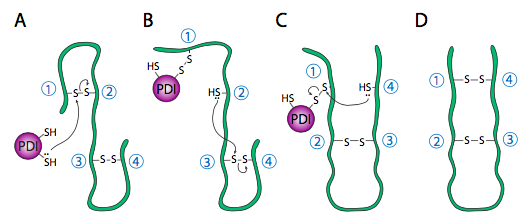
El replegamiento de proteínas es un proceso importante porque los patrones de plegamiento iniciales como el polipéptido aún se están traduciendo y sin terminar puede no ser el patrón de plegamiento óptimo una vez que la proteína completa está disponible. Esto es cierto no solo de los enlaces H, sino también de los enlaces disulfuro más permanentes (es decir, covalentes). Al observar el hipotético ejemplo de polipéptido, la estructura secundaria de la mitad N-terminal puede conducir a la formación de un enlace disulfuro estable entre la primera cisteína y la segunda cisteína, pero en el contexto de la proteína completa, se podría formar un enlace disulfuro más estable entre la cisteína 1 y cisteína 4. El intercambio de dianas de enlaces disulfuro es catalizado por la proteína disulfuro isomerasa (PDI).
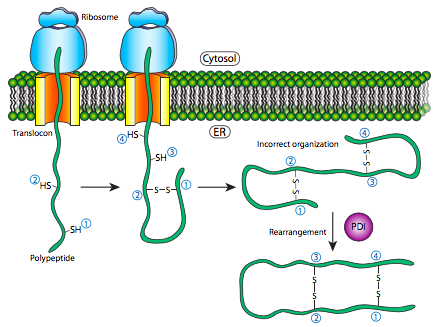
El ambiente redox interno del retículo endoplásmico, es significativamente más oxidativo que el del citoplasma. Esto está determinado en gran medida por el glutatión, el cual se encuentra en una relación de 30:1 GSH:GSSG o superior en el citoplasma pero a una relación casi 1:1 en la luz del ER. Este ambiente oxidativo también es propicio para la remodelación de disulfuro. Cabe señalar que PDI no elige a los socios de vinculación “correctos”. Simplemente mueve los enlaces disulfuro existentes a una disposición más energéticamente estable. A medida que el resto del polipéptido continúa replegándose, rompiendo y haciendo enlaces H rápidamente, nuevos socios potenciales de enlaces disulfuro pueden moverse cerca unos de otros y PDI puede intentar nuevamente reorganizar el patrón de enlaces disulfuro si el patrón resultante es más termodinámicamente estable.
El ensamblaje de proteínas multisubunitarias y el replegamiento de polipéptidos son similares en su uso de proteínas chaperonas que ayudan a prevenir el plegamiento prematuro, secuestrando partes de la proteína de la interacción de enlace H hasta que la proteína completa esté en la luz del RE.
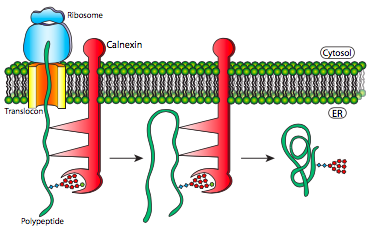
Este mecanismo simplemente facilita la búsqueda de la conformación termodinámicamente óptima al evitar la formación de algunas conformaciones subóptimas potenciales. Estas proteínas chaperonas se unen a las nuevas proteínas a medida que ingresan al lumen a través del translocon y además de simplemente prevenir enlaces incorrectos que tendrían que romperse, también evitan la interacción prematura de múltiples polipéptidos entre sí. Esto puede ser un problema porque antes del plegamiento adecuado que normalmente ocultaría dichos dominios dentro de la proteína, los polipéptidos inmaduros pueden tener dominios de interacción expuestos, conduciendo a una unión indiscriminada, y potencialmente precipitación de agregados de proteínas insolubles.
Chaperones
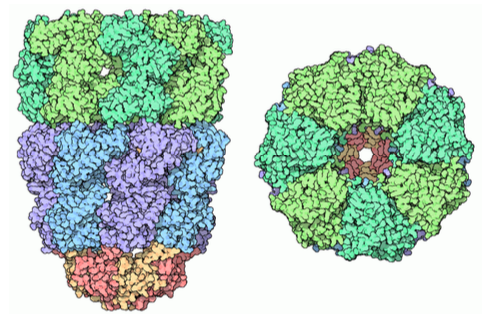
Las proteínas chaperonas también se pueden encontrar en procariotas, arqueas y en el citoplasma de eucariotas. Estas son algo similares entre sí, y funcionan de manera algo diferente a los tipos de proteínas de plegamiento que se encuentran en la luz del RE. Se les conoce generalmente como chaperoninas, y el mejor caracterizado es el complejo Groel/ GROEs en E. coli. Como\(\PageIndex{11}\) indica la estructura en la Figura, es similar en forma al proteasoma, aunque con una función completamente diferente. GroEL se compone de dos anillos apilados, cada uno compuesto por 7 subunidades, con una gran cavidad central y una gran área de residuos hidrófobos en su apertura. GROEs también se compone de 7 subunidades, y actúa como una tapa en un extremo del GroEl. Sin embargo, los GROEs solo rematan a GroEl en presencia de ATP. Tras la hidrólisis del ATP, las chaperoninas experimentan importantes cambios conformacionales concertados que inciden en la proteína en su interior, provocando el replegamiento, y luego los GROEs se disocian y la proteína se libera de nuevo al citosol.