
12.1: Introducción al citoesqueleto
- Page ID
- 53346

Cuando una célula eucariota se saca de su contexto fisiológico y se coloca en una placa de Petri de plástico o vidrio, generalmente se ve que se aplana hasta cierto punto. En un precipicio, se comportaría como un reloj Salvador Dalí, rezumando por encima del borde. La suposición inmediata, particularmente a la luz de que se sabe que la celda es mayormente agua en masa y volumen, es que la celda es simplemente una bolsa de fluido. Sin embargo, la célula en realidad tiene una intrincada microestructura dentro de ella, enmarcada internamente por los componentes del citoesqueleto.
Aunque los genes no están particularmente bien conservados, una combinación de similitud genética y estructura proteica han confirmado la presencia de proteínas procariotas que están relacionadas con proteínas citoesqueléticas eucariotas tanto en forma como en función. En comparación con el citoesqueleto eucariota, el estudio de las proteínas procariotas es muy reciente, y durante mucho tiempo, se asumió que los procariotas no tenían o necesitaban arquitectura citoesquelética. FtsZ, el equivalente bacteriano de la tubulina, fue descubierto en 1980 pero la mayor parte del trabajo en él se ha producido en la última década. MReB es una proteína similar a actina, primero en comparación con la actina en 1992, y la crescentina, una proteína de clase de filamentos intermedios, solo se describió en 2003. Para una revisión integral de las proteínas del citoesqueleto procariota, véase Graumann, P.L., Ann. Rev. Microbiología 61:589-618, 2007.
Como su nombre lo indica, el citoesqueleto actúa de manera muy similar a nuestros propios esqueletos al sostener la forma general de una célula. Sin embargo, a diferencia de nuestros esqueletos, el citoesqueleto es altamente dinámico e internamente móvil, desplazándose y reordenándose en respuesta a las necesidades de la célula. También tiene una variedad de propósitos más allá de simplemente proporcionar la forma de la célula. Generalmente, estos pueden ser categorizados como estructurales y de transporte. Si bien los tres componentes principales del citoesqueleto realizan cada una de estas funciones, no lo hacen por igual, ya que sus características biofísicas son bastante diferentes. Con respecto a la estructura, en algún momento de la vida de cada célula, debe cambiar de forma, ya sea simplemente aumentando o disminuyendo de tamaño, o una alteración más drástica como la forma súper alargada de neuronas con axones, el citoesqueleto debe ser capaz de responder aumentando y disminuyendo dinámicamente el tamaño de las estructuras internas según sea necesario. La estructura también se aplica a la posición relativa de elementos celulares internos, como orgánulos o proteínas, entre sí. En muchas células altamente especializadas, la segregación de estructuras particulares dentro de ciertas partes de la célula es crucial para que funcione. El transporte se refiere al movimiento de moléculas y orgánulos dentro de la célula así como al movimiento de la célula en su conjunto. Acabamos de discutir el movimiento intracelular de proteínas y lípidos a través de vesículas en el último capítulo. Esas vesículas, como veremos en este capítulo, no solo están flotando de un lugar a otro; se mueven a propósito y direccionalmente a lo largo del citoesqueleto como carga en carreteras o vías férreas. Con respecto al movimiento de células enteras, esto puede ir desde remar o nadar por organismos unicelulares hasta el rastreo estereotipado y altamente coordinado de muchas células desde su punto de origen hasta su destino final durante el desarrollo de un organismo metazoico o el movimiento de fibroblastos para curar un corte en tu piel.
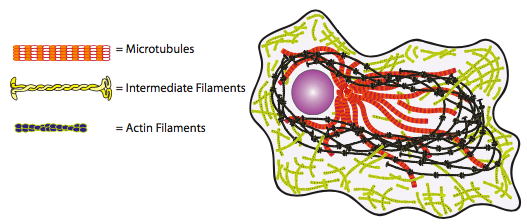
Los tres componentes principales del citoesqueleto son microtúbulos, microfilamentos y filamentos intermedios. Cada uno de estos son polímeros compuestos por subunidades repetitivas en arreglos específicos. Con solo un vistazo rápido (Figura\(\PageIndex{1}\)), es muy claro que los filamentos intermedios probablemente jugarán un papel significativamente diferente de los microtúbulos o microfilamentos. Debido a que los IF están hechos de subunidades fibrosas largas que se enrollan unas alrededor de otras para formar el filamento, claramente existe un gran contacto (lo que facilita la formación de enlaces de hidrógeno, también conocido como velcro molecular TM) entre las subunidades proporcionando una gran resistencia a la tracción. Es muy difícil separar estas subunidades y, por lo tanto, los IF se utilizan principalmente para fines de soporte de carga permanente o a largo plazo. Al observar los otros dos componentes del citoesqueleto, se puede ver que con la forma globular en lugar de fibrosa de las subunidades, el área máxima de contacto entre las subunidades es muy limitada (piense en el área de contacto cuando empuja dos balones de baloncesto juntos), facilitando la separación de las subunidades o romper el microfilamento o microtúbulo. La célula puede aprovechar esta característica a su favor, al utilizar este tipo de fibras citoesqueléticas en situaciones dinámicas donde la formación o destrucción de filamentos intermedios tardaría demasiado tiempo. Ahora abordamos estos tres grupos de elementos citoesqueléticos con más detalle.