
15.6: Mitosis
- Page ID
- 53291

La mitosis consiste en profase, metafase, anafase y telofase, con distintas actividades celulares que caracterizan cada fase. Esto completa la duplicación del núcleo, y es seguida por la citocinesis, en la que la célula se divide para producir dos células hijas.
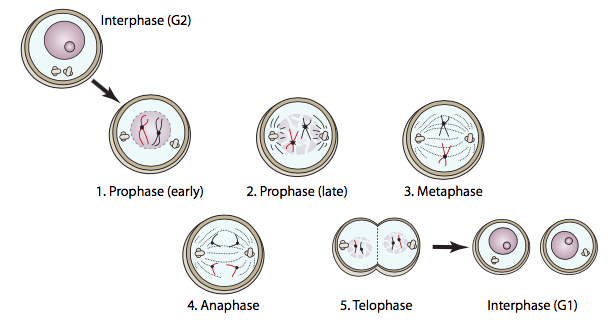
La profase es la preparación de cada componente para esta compleja danza celular. El ADN se condensa (se envuelve alrededor de sí mismo firmemente para convertirlo en un paquete más pequeño y más fuerte) de manera que es menos susceptible a la rotura durante el movimiento a través de la célula. Al hacerlo, la mayor parte del ADN se vuelve transcripcionalmente inactivo. Los cuerpos de Golgi y el retículo endoplásmico comienzan a romperse en vesículas membranosas que pueden distribuirse de manera más fácil y uniforme a través de la célula para que ambas células hijas reciban aproximadamente lo mismo. Los centrosomas (en células animales) se mueven desde su posición original cerca del núcleo hacia lados opuestos de la célula, para establecer los polos del huso mitótico.
El MPF fosforila las proteínas motoras de los microtúbulos y las proteínas asociadas a microtúbulos (MAP) para alterar la dinámica normal de los microtúbulos y permitir que se produzca la reorganización masiva en un huso mitótico. Por ejemplo, una diana de MPF es PRC1, una proteína de agrupación que se inactiva por fosforilación, permitiendo así que los microtúbulos individuales se muevan a nuevas ubicaciones más fácilmente de lo que podría hacer un haz grande. Otros efectos son la inactivación de MAP estabilizantes, lo que conduce a una mayor labilidad de los microtúbulos debido al aumento de las incidencias de catástrofe. Las proteínas motoras dianas de MPF están en la familia de la cinesina y la fosforilación es necesaria para que algunas de ellas se unan al huso mitótico.
La prometafase a veces se considera una fase separada, pero también se conoce como profase tardía, y se define principalmente por la ruptura de la envoltura nuclear. Este proceso es inducido por la fosforilación de MPF de las laminas nucleares. Adornadas con cargas negativas de los fosfatos, las laminas se niegan a asociarse entre sí por más tiempo, lo que lleva a la descomposición de la lámina nuclear. A medida que las laminas se disocian, la envoltura nuclear permanece unida a ellas, y fragmentos. Esta fragmentación nuclear debe ocurrir para que el huso mitótico pueda llegar al interior y adherirse a los cromosomas.
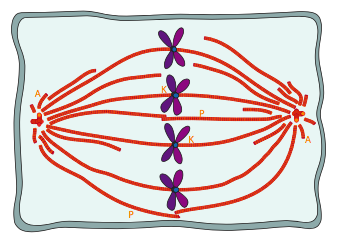
Algunos de los microtúbulos del huso mitótico se unen a los cromosomas a través de las proteínas cinetocoras, las cuales enlazan los microtúbulos del huso con la región del centrómero de cada cromosoma. Estos se conocen como microtúbulos de cinetocoros (Figura\(\PageIndex{8}\)). Existen otros dos tipos de microtúbulos en el huso mitótico (Figura\(\PageIndex{7}\)): los microtúbulos polares que llegan a través de la célula e interactúan entre sí para ayudar a mantener la separación de los centrosomas y definir la longitud total del huso, y los microtúbulos aster que generalmente son corto, irradiando y estabilizando el centrosoma. Recuerde que el ADN replicado más temprano en la fase S, y así las cromátidas hermanas todavía están parcialmente unidas. Visualmente, la región del centrómero parece más estrecha o más comprimida que el resto del cromosoma, y generalmente se encuentra cerca del centro. El centrómero contiene secuencias repetidas que están involucradas en la unión y ensamblaje de cinetocoros.
En los primates, el motivo de repetición se conoce como ADN satélite alfa, que está compuesto por múltiples instancias de repeticiones en tándem de una secuencia central de ~170 pb sobre un tramo de ADN centromérico sobre una megabasa de longitud. También se encuentran repeticiones similares en otros vertebrados. En otros eucariotas, el tamaño y la secuencia pueden variar; por ejemplo, se encuentran repeticiones mucho más cortas de ~5 pb en el ADN centromérico que mide 200-600kb en cromosomas de Drosophila, y S. pombe tiene ADN centromérico muy por debajo de 10kb.
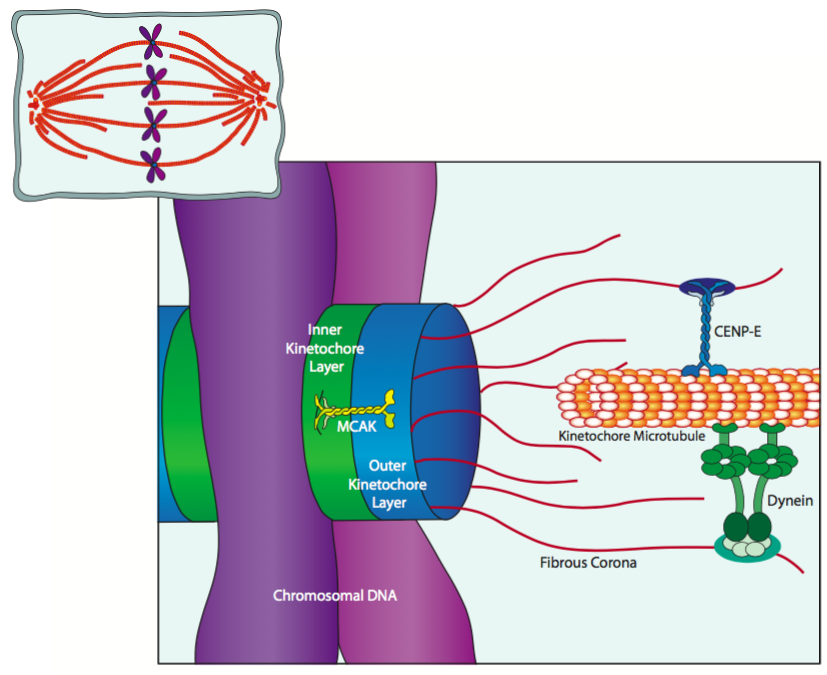
Los cinetocoros que se unen al ADN del centrómero son estructuras proteicas trilaminares que consisten en una capa interna, una capa externa y una corona fibrosa. Los microtúbulos cinetocoros del huso mitótico se unen principalmente a la corona fibrosa. Como se representa en la Figura, se une a través de CENP-E, una cinesina y proteínas motoras de dineína que se unen a lo largo del barril del microtúbulo. De hecho, a veces el primer contacto entre un cromosoma (a través del cinetocoro) y un microtúbulo huso se encuentra en algún lugar en el medio del microtúbulo, y una combinación de dinámica de microtúbulos y actividad de proteínas motoras mueven el cromosoma al extremo distal del microtúbulo. Esto es facilitado por MCAK (cinesina asociada al centromero mitótico), que se asocia con las proteínas del núcleo del cinetocoro y juega un papel en la despolimerización de microtúbulos cerca del extremo (+).
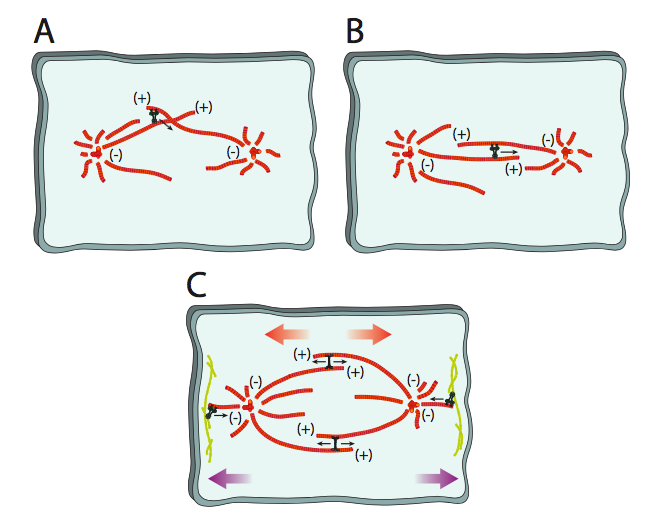
La transición del citoesqueleto de microtúbulos interfásicos a un huso mitótico requiere una serie de motores moleculares para mover los centrosomas, alinear los microtúbulos y expandir el huso. Estos se representan en la Figura\(\PageIndex{9}\). Inicialmente, a medida que los centrosomas duplicados se alejan entre sí junto con algunos de los microtúbulos citoesqueléticos, los microtúbulos interactuarán en varios ángulos. Debido a que los microtúbulos polares que ayudan a expandir o mantener el ancho del huso deben ineractuar en paralelo, las dineinas citoplásmicas se unen a los eventuales microtúbulos polares y al moverse uno a lo largo del otro, los llevan en paralelo (9a). Una vez en esa posición, BiMC y otras quinesinas toman el relevo como motores primarios a lo largo de los microtúbulos polares. Crean una fuerza de empuje hacia afuera al sujetarse a un microtúbulo orientado hacia una dirección y conducir a lo largo de un MT paralelo orientado hacia la dirección opuesta hacia el extremo (+) (9b). Finalmente, las dineinas citosólicas unidas al citoesqueleto cortical tiran de los microtúbulos astrales, lo que tira de los extremos del huso más lejos del centro (9c).
De hecho, parece haber dos mecanismos en funcionamiento: el sistema bub1/bub2 funciona en la vía de detección de tensión, mientras que otra proteína metafásica, mad2, parece ser importante en la suspensión de la mitosis tras la desconexión del cinetocoro con el microtúbulo del huso.
A medida que la envoltura nuclear se está rompiendo, los microtúbulos del huso mitótico están experimentando una mayor inestabilidad dinámica, ciclando entre períodos de brotes de crecimiento (polimerización) y acortamiento rápido (desmontaje catastrófico), buscando cromosomas a los que conectarse. Una vez que los microtúbulos del cinetocoro se conectan a los cromosomas, la dinámica de los microtúbulos cambia. El microtúbulo sufrirá principalmente un acortamiento si está más allá del centro del husillo y principalmente se alargará si está corto de centro. Dado que eventualmente cada conjunto de cromátidas hermanas está conectado a microtúbulos en ambos cinetocoros, cada cromátida está conectada a un microtúbulo de acortamiento y un microtúbulo de alargamiento. A medida que los cromosomas se acercan al centro del huso mitótico, la tasa de acortamiento/alargamiento de los microtúbulos se ralentiza. Las cromátidas hermanas son empujadas y tiradas por los microtúbulos del huso hasta que se alinean a lo largo de la línea media del huso mitótico, que en la mayoría (pero no en todos) los casos es también la línea media de la célula. Una vez que están todos alineados, se considera que la célula ha alcanzado la metafase. A diferencia de las otras fases, la metafase es una fase relativamente estática, es un punto de control para alinear los cromosomas.
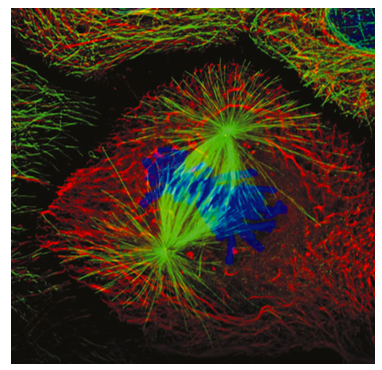
Los cromosomas deben estar correctamente alineados para asegurar que ambas células hijas reciban el complemento adecuado de los cromosomas. ¿Cómo sabe la célula cuándo los cromosomas han llegado al centro del huso? Un experimento elegantemente simple demostró que el mecanismo general es un control de tensión: si los dos microtúbulos que se conectan al par de cromátidas hermanas de cada lado son de la misma longitud, deberían estar ejerciendo la misma tensión sobre los cromosomas. Si la conexión microtúbulo-cinetocoro se corta en la metafase, se evitará que la célula progrese (Nicklas, R.B., et al, J. Cell Biol. 130: 929-39, 1995). Sin embargo, si se aplica una tensión equivalente tirando del cromosoma con una microaguja de vidrio, ¡se restablece la progresión de la mitosis!
Además del chequeo de tensión, hay otra condición que debe cumplirse para continuar con la mitosis: el MPF debe ser inactivado. Como se indicó anteriormente, MPF en parte conduce a su propia inactivación al activar el complejo promotor de la anafasa (APC), que poliubiquitina la ciclina, conduciendo a su destrucción y por lo tanto a la inactivación de MPF-CDK. APC también etiqueta securin para su destrucción. La securin es una proteína que se une e inhibe la enzima proteolítica, la separasa, cuya activación es necesaria para permitir que las cromátidas hermanas se separen, lo que a su vez, es necesario para que proceda la anafase.
Salvo situaciones patológicas, si y sólo si todos los cromosomas se alinean en la placa de metafase, la célula procederá a la siguiente etapa de mitosis: la anafase. Las cromátidas hermanas se separan y son tiradas hacia polos opuestos del huso mitótico. Algo perversamente, aun cuando los cromosomas se mueven hacia los polos del huso, los polos mismos se mueven ligeramente hacia afuera. La separación de las cromátidas hermanas requiere la disociación del “pegamento” molecular que las mantiene unidas: las proteínas cohesina. Las cohesinas se unen a ambas moléculas de ADN y las mantienen unidas poco después de la replicación en la fase S. A medida que se acerca la anafase, se activa la enzima separasa, que luego corta las moléculas de cohesina. Una vez cortadas todas las moléculas de cohesina, las cromátidas hermanas finalmente se pueden separar. La remoción de las cohesinas procede aproximadamente hacia adentro desde los puntos distales de los cromosomas hasta el centrómero, que generalmente es la última región de unión.
Una cohesina es un multímero de cuatro subunidades, Sc1, Sc3, Smc1 y Smc3 en levadura. También se ha observado una proteína adicional en Xenopus. La proteína SCC1 se escinde por separina en levaduras, pero en metazoos, la SCC1 también se puede eliminar de los cromosomas por otro método. Se fosforila, lo que disminuye su afinidad por el ADN, y puede exponer un sitio para hidrólisis catalizada por separación.
La separasa también promueve la anafase activando Cdc14, una fosfatasa necesaria para desfosforilar los sustratos cdk que habían sido fosforilados por los complejos ciclina-cdk de mitosis temprana. Además, también se requiere Cdc14 para citocinesis en la levadura S. cerevisiae y el nematodo C. elegans.
La anafase se puede dividir en dos etapas, algunas veces denominadas temprana y tardía o A y B. Al principio, los microtúbulos de cinetocoros se acortan desde ambos extremos, y los motores de la familia kinesina tiran de los microtúbulos hacia los polos del husillo. A medida que comienza la anafase tardía, los microtúbulos polares se alargan y una fuerza adicional de separación cromátida es aplicada por las proteínas motoras de la familia quinesina [quinesina-5] que empujan los microtúbulos polares entre sí para aumentar la separación entre los polos. Los motores de la familia dineína también ayudan a dirigir el movimiento de los polos, a través de su unión a los microtúbulos aster y al citoesqueleto cortical (periférico).
Cuando ambos conjuntos de cromosomas llegan a sus respectivos polos, comienza la telofase. Técnicamente, se estaba acumulando lentamente desde la anafase: cuando el MPF fue inactivado por APC, su capacidad para fosforilar laminas nucleares terminó. Las proteínas fosfatasas en la célula eliminan los grupos fosfato, permitiendo que las laminas interactúen una vez más entre sí, y por telofase están reconstituyendo la lámina nuclear y la envoltura nuclear. Dado que las laminas y otras proteínas de la membrana nuclear también interactúan con el ADN, los fragmentos de membrana nuclear dispersados en la profase tardía ahora se fusionan alrededor de cada conjunto de ADN para formar las nuevas envolturas nucleares. Los otros orgánulos membranosos fragmentados (ER, Golgi) también comienzan a reformarse. Al final de la telofase, el producto es una sola célula grande con dos núcleos completos en lados opuestos. El siguiente y último paso, la citocinesis, divide la célula en dos células hijas separadas e independientes. En las células animales, la citocinesis es similar al apriete de un cordón en el centro de la célula, tirando de la “cintura” hasta que todos los bordes se encuentran, y se obtienen dos células separadas. Este anillo contráctil está compuesto por subunidades actina (estructural) y miosina (motivo). Estas proteínas, que utilizan ATP para obtener energía, se encadenan cada vez más cerca de manera similar al “golpe de potencia” de actina y miosina descrito para los sarcómeros de células musculares, también hechas principalmente de actina y miosina. Este mecanismo es universal para las células animales, pero la colocación del anillo no siempre está en el centro de la célula. El anillo suele coincidir con el centro de la célula, pero de hecho está posicionado por la placa metafásica (es decir, el centro del huso mitótico). El ejemplo más obvio de una placa metafásica que no coincide con el centro de la célula se encuentra en la formación de óvulos. Debido a que el propósito de un óvulo es proporcionar todo el material necesario para hacer un nuevo organismo viable tras la fertilización (el esperma aporta biomasa insignificante más allá del material genético), se divide asimétricamente, con el huso mitótico localizado lejos a un lado de la célula (Figura\(\PageIndex{11}\)).
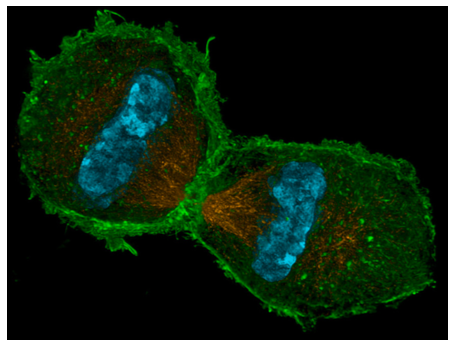
Cuando ocurre la citocinesis, una célula hija, el presunto ovocito, es muy grande, mientras que la otra célula, llamada cuerpo polar, tiene mínimo material citoplásmico que rodea el núcleo. El anillo contráctil funciona en las células animales porque la membrana celular es flexible. En las células vegetales, la membrana celular está firmemente unida a una pared celular rígida y, por lo tanto, no se puede introducir. Entonces, la célula vegetal construye ingeniosamente una pared por la mitad de la célula utilizando vesículas especializadas que se originan en parte del Golgi, y que contienen los materiales necesarios para formar una pared celular. Las vesículas viajan a lo largo del phragmoplast, una estructura construida a partir de los microtúbulos del huso mitótico, y a medida que las vesículas se alinean a lo largo de la mitad de la célula, comienzan a fusionarse para formar vesículas más grandes y luego una vesícula grande en forma de disco, la placa celular. Eventualmente llegan a la membrana celular misma, y fusionándose con eso conduce a la formación de una nueva pared celular, y dos células completas e independientes.
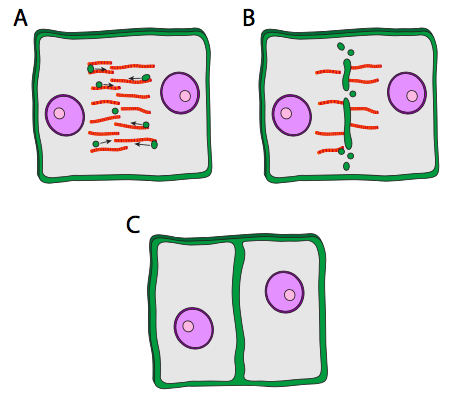
El contenido de las vesículas que viajan a lo largo del phragmoplast no está bien descrito. Se sabe que la calosa, un polisacárido de glucosa con enlaces β1-3, está presente en la placa celular en desarrollo, pero no se ha encontrado en el Golgi ni en las vesículas. Curiosamente, una vez que la placa celular se ha fusionado completamente con las paredes celulares existentes, la calosa desaparece gradualmente. Se piensa que el mismo sistema enzimático que sintetizó la calosa puede cambiar a sintetizar celulosa a medida que la placa celular madura.