
6.4: Análisis genéticos de la biosíntesis de metionina
- Page ID
- 53602

Al observar la vía para la biosíntesis del Met más adelante en este capítulo, puede preguntarse cómo se asociaron los números de genes con genes específicos, ya que los números no corresponden a las posiciones de las reacciones codificadas por los productos génicos MET en la ruta. El sistema de numeración refleja el proceso de descubrimiento de los genes MET. Los primeros estudios de biosíntesis de Met
en levaduras fueron realizados por genetistas, quienes utilizaron cribas genéticas clásicas para aislar mutantes met. Las pantallas genéticas son herramientas importantes para identificar nuevos genes porque son imparciales por el conocimiento previo de la vía. Además, la mutación es un proceso aleatorio que debe afectar a todos los genes involucrados en la producción del fenotipo en estudio. El genetista comienza tratando una cepa parental con una sustancia química o radiación para inducir mutaciones en el ADN. La tasa de mutación espontánea en
levaduras es de ~10-8/base/generación, que es demasiado baja para un cribado genético práctico. Por lo tanto, los investigadores utilizan dosis mutágenas que matan hasta ~ 50% de las células. Las células que sobreviven a la mutagénesis suelen albergar un gran número de mutaciones, muchas de las cuales no tienen ningún efecto sobre el fenotipo
que se está explorando. En consecuencia, se requieren grandes cantidades de células para descubrir todos los genes involucrados en el fenotipo. Por ejemplo, el genoma de la levadura contiene ~6000 genes, por lo que un cribado genético útil podría involucrar 20,000 o más células.
Los medios selectivos proporcionan herramientas importantes para identificar fenotipos mutantes en pantallas genéticas. Dependiendo del fenotipo que se esté estudiando, los investigadores pueden seleccionar mutantes usando un esquema de selección positivo o negativo, como se muestra en la página opuesta. Los tipos de cribado más fáciles emplean selección positiva, ya que solo las células mutantes crecen en medios selectivos. Si los investigadores están analizando vías que son importantes para el crecimiento celular, como la síntesis de Met, probablemente usarían un esquema de selección negativa. En un esquema negativo, las células se cultivan primero
Estrategias de selección utilizadas para aislar mutantes de levadura.
Después de la mutagénesis inicial, las levaduras se cultivan en una placa que contiene medios ricos (o sintéticos completos). En esta figura, la mutagénesis ha generado tres mutantes diferentes en el gen de interés. Las colonias mutantes
están rodeadas por un círculo vacío. Las réplicas de la placa maestra se copian en medios selectivos. En un esquema de selección negativa, la placa selectiva carece de un componente que normalmente está presente en medios ricos. En un esquema de selección positiva, el medio contiene un agente selectivo, que es tóxico para las células normales, pero tolerado por las células mutantes. El agente selectivo es a veces un análogo tóxico de un metabolito celular normal.
en medios, como YPD o YC, que permiten que todas las células crezcan. Las réplicas de estas placas maestras se realizan luego en medios definidos que carecen de Met. (El chapado de réplica se describe en el Capítulo 12.) Dado que solo las células de tipo silvestre crecen en los medios selectivos que carecen de Met, los investigadores buscan colonias en los medios ricos cuyas contrapartes faltan en los medios selectivos.
El número y espectro de mutantes obtenidos en un cribado genético son impredecibles, debido a la naturaleza aleatoria de la mutación. Como cabría esperar, una pantalla podría producir
múltiples mutantes en un gen y ninguna mutación en otros genes involucrados en el fenotipo.
Después de completar un cribado, los investigadores deben determinar a continuación si las mutaciones están en el
mismo o en diferentes genes. Para ello, los genetistas se basan en el mapeo genético (Capítulo 5) y/o la complementación. La complementación es una prueba funcional de la actividad génica. En un experimento de complementación, la introducción de un gen funcional de otra fuente rescata un fenotipo mutante causado por el gen defectuoso. La complementación genética clásica en levaduras aprovecha los
dos tipos de apareamiento de levaduras y la capacidad de la levadura para sobrevivir como cepas tanto haploides como diploides. En un experimento de complementación con mutantes met, los investigadores aparean un mutante met haploide en el tipo de apareamiento a o a (MAT a o MAT a) con un mutante met haploide del tipo de apareamiento opuesto. Si el diploide es capaz de crecer en ausencia de Met, se ha producido la complementación, y las mutaciones met en las dos cepas haploides deben estar en genes diferentes. Si el diploide no es capaz de sobrevivir en la placa selectiva, las dos cepas haploides portan mutaciones en el mismo gen (aunque es casi seguro que son alelos mutantes diferentes). Un cribado genético puede producir múltiples alelos mutantes del mismo gen, que juntos forman un grupo de complementación.
Para 1975, los laboratorios de levaduras habían aislado colecciones de mutantes met y mapearon nueve de las mutaciones met en cromosomas. En un estudio histórico, Masselot y Derobichon-szulmajster (1975) recolectaron 100 cepas met de laboratorios de todo el mundo e hicieron experimentos sistemáticos de complementación con todos los mutantes. Se identificaron 21 grupos de complementación, que representan genes potenciales, y a los genes se les asignaron nombres MET1 a MET25. Muchos de los genes MET codifican enzimas en la ruta biosintética de Met, que se describe en la página opuesta. Algunos productos génicos están involucrados en la síntesis de cofactores y donantes de metilo utilizados
en la ruta, mientras que otros productos génicos MET (no mostrados) están involucrados en la regulación de la vía (revisado en Thomas & Surdin-Kerjan, 1992). En su mayor parte, los nombres asignados en el estudio de 1975 todavía se utilizan en la actualidad. Posteriormente se demostró que algunos genes identificados en el estudio de 1975 no estaban involucrados en la biosíntesis de Met, y otros (por ejemplo, MET15, MET17 y MET25) representaban diferentes alelos del mismo gen (D'Andrea et al., 1987).
En la época del estudio de 1975, las reacciones bioquímicas en la vía eran ampliamente conocidas, y los científicos enfrentaron el desafío de asociar genes con actividades enzimáticas. Se puede ver por la vía que las mutaciones en 11 genes MET diferentes producirían un fenotipo en el que las cepas crecerían en presencia de metionina, pero no en su ausencia. Los científicos estrecharon posibles relaciones geno-enzima analizando la capacidad de las cepas met para
utilizar fuentes alternativas de azufre en lugar de metionina (Masselot y Derobichon-szulmajster, 1975). Las levaduras son muy versátiles en su uso de fuentes de azufre tanto inorgánicas como orgánicas. El sulfato es transportado eficientemente a las células por los transportadores Sul1p y Sul2p en la membrana. El sulfito y el sulfuro también se transportan a las celdas con una eficiencia reducida. Las levaduras también son capaces de transportar y utilizar Met, Cys, homocisteína y S-adenosilmetionina (AdoMet o SAM) como fuentes de azufre (revisado en Thomas y Surdin-Kerjan, 1992). En este laboratorio, utilizará medios selectivos en los que el sulfito o la cisteína reemplazan a la metionina para distinguir entre 3 mutantes met. También se utilizará un medio diferencial, el agar BigGy, que distingue a las cepas de levadura por su producción de sulfuro de hidrógeno. Los medios diferenciales permiten que todos los mutantes crezcan, pero los mutantes producen colonias que pueden distinguirse entre sí por su color o morfología.
Los mutantes met utilizados en este curso NO fueron generados por mutagénesis tradicional. En cambio, los mutantes fueron construidos por un enfoque molecular más nuevo que requiere un conocimiento detallado de la secuencia del genoma de la levadura. Después de que se completó el proyecto del genoma de levadura, los investigadores se interesaron en obtener una colección de cepas de deleción en todo el genoma, cada una de las cuales difería de la cepa BY4742 parental en un solo locus génico. Su enfoque, que
se discute con más detalle en el Capítulo 7, aprovecha la alta frecuencia con la que S. cerevisiae se somete a recombinación homóloga. Cada ORF en el genoma de S. cerevisiae se reemplazó sistemáticamente por un gen bacteriano KAN R (Winzeler et al., 1999). Una ventaja importante de esta estrategia, a veces denominada “genética inversa”, sobre el enfoque genético tradicional es que la selección positiva puede usarse para aislar mutantes. Solo las cepas con genes MET alterados son capaces de crecer en medios que contienen análogos de kanamicina. Las cepas con genes KAN R alterados tienen otras ventajas sobre las cepas mutantes generadas con mutágenos químicos o tratamiento con radiación.
Las cepas no albergan mutaciones secundarias inducidas por el tratamiento mutágeno y no es posible la reversión espontánea a un fenotipo silvestre.
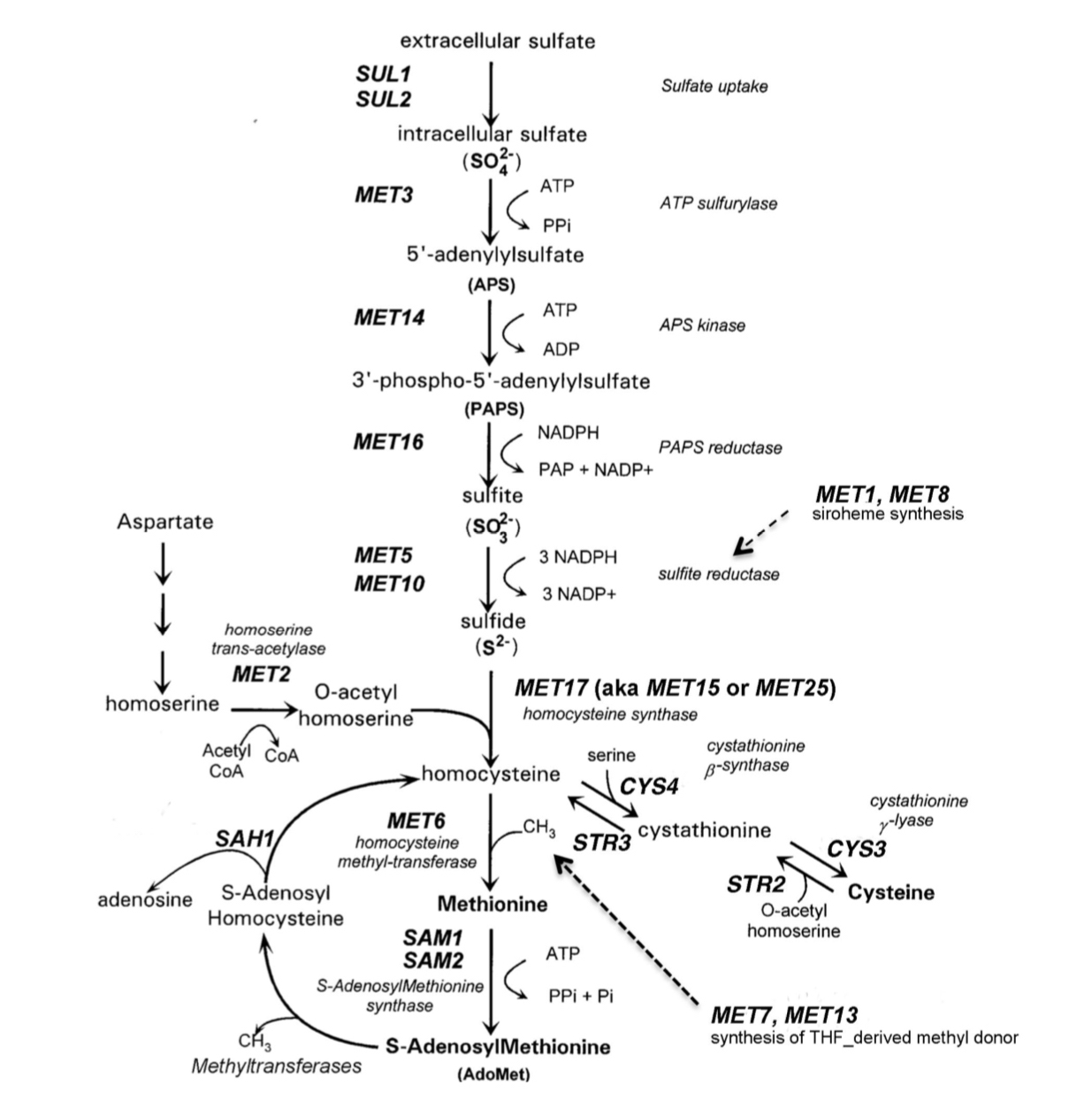
Biosíntesis de metionina en levaduras.
Las proteínas que catalizan etapas individuales en la biosíntesis de Met y Cys se enumeran junto a cada paso de la ruta. Los nombres de los genes que codifican las actividades se muestran en mayúsculas en cursivas, siguiendo las convenciones de
S. cerevisiae. Los genes MET1 y MET8 codifican proteínas que participan en la síntesis del sirohemo, un cofactor esencial para la sulfito reductasa. Los productos génicos MET7 y MET13 catalizan los dos últimos pasos en la síntesis del donante de metilo utilizado por Met6p, homocisteína metiltransferasa, para sintetizar metionina. (Adaptado de Thomas et al. ,1992)