5.1: Desdoblando el eje A/P: Genes Principio-Hox, Otro Nivel de Regionalización
- Page ID
- 53009

Regionalización a Gran Escala: Genes Hox y la Cascada de Segmentación
Ya echamos un vistazo rápido a los genes Hox, tanto en tus lecturas como en la sección Genetic Toolkit. Ahora los pondremos en un contexto más amplio de Evo-Devo observando un ejemplo específico de patrones de Hox (dando seguimiento a la historia El curioso caso del bicoide) y al ver evidencias de que la regionalización por genes Hox se conserva a través de los animales. También se conservan algunos elementos cis-reguladores que impulsan los patrones de expresión del gen Hox. Sin embargo, los eventos de desarrollo anteriores que establecieron dominios de expresión de Hox no se conservan entre los animales, difieren ampliamente a veces incluso dentro de un filo. Por ejemplo, aunque todos los artrópodos están segmentados en Drosophila, los segmentos se modelan simultáneamente, mientras que en algunos otros insectos y artrópodos surgen uno a la vez. La conservación del patrón Hox a pesar de la variabilidad de la generación más amplia de patrones A/P sugiere que el patrón Hox representa una restricción del desarrollo en la evolución animal. Es decir, una vez que evolucionó el patrón Hox, fue tan ventajoso que en su mayoría se conservó durante millones de años de evolución animal.
Nos centramos en Drosophila a pesar de que es un caso extraño de regionalización simultánea porque está muy bien estudiada y más fácil de entender que muchos otros organismos modelo. Nuestra mirada a la regionalización y el patrón A/P de Drosophila ilustrará tres conceptos principales en Evo-Devo:
- Hay múltiples formas de llegar a etapas de desarrollo restringidas.
- Gran parte del desarrollo ocurre dividiendo repetidamente tejidos/regiones y especificándolos en destinos más restringidos.
- Múltiples salidas de expresión génica pueden superponerse entre sí para dar a una célula/tejido su identidad específica.
En la escisión y gastrulación, observamos la idea de que la simetría se rompe en la creación de ejes y capas germinales en animales. También vimos que los gradientes de proteína Bicoid y Nanos opuestos son capaces de especificar la primera regionalización anteroposterior en el desarrollo de moscas de la fruta. Pero si pensamos en una mosca adulta, tienen muchas más regiones anteroposteriores que simplemente una cabeza y una cola. Por ejemplo, sólo algunos segmentos llevan alas o piernas, otros llevan ojos o antenas y otros tienen genitales u órganos excretores. ¿Cómo se “regionaliza” la mosca en desarrollo, es decir, pasar de tener dos gradientes anteroposteriores opuestos a tener patrones anteroposteriores a pequeña escala? Para responder a esto, veremos la genética del desarrollo de la cascada de segmentación, la serie de pasos que divide a la mosca embrionaria en subdivisiones cada vez más finas.
La cascada de segmentación de Drosophila
Aquí voy a resumir muy brevemente la cascada de segmentación, para más detalles por favor visite Los orígenes de la polaridad anterior-posterior
La cascada de segmentación es jerárquica tanto en el tiempo de desarrollo como en la regulación de la expresión génica. Los genes de efecto materno se expresan más temprano y regulan los genes gap. Los genes Gap se expresan a continuación, actuando con los genes de efecto materno para regular los genes de regla de pares. Los genes de regla de pares y genes gap regulan la expresión de los genes segmento-polaridad, los cuales se expresan en último lugar (Figura 1).
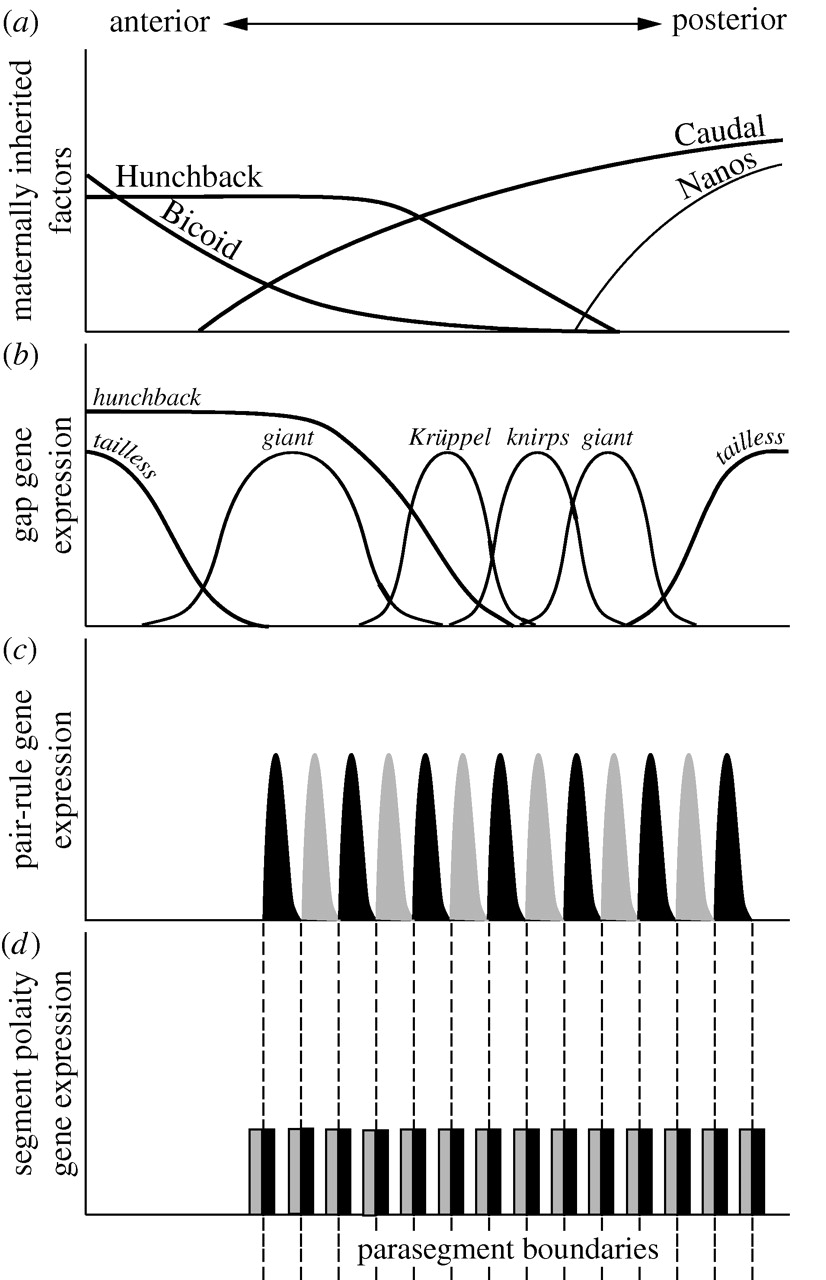 |
Figura 1: La cascada de segmentación de Drosophila. Drosophila divide todos sus segmentos a la vez, pero la mayoría de los artrópodos (un animal en general) crecen a partir de una zona de crecimiento posterior donde las regiones más anteriores se desarrollan primero. Considere qué partes de esta jerarquía de expresión y especificación podrían conservarse a través de diferentes modos de desarrollo y cuáles partes no podrían ser. De “La evolución de las redes génicas del desarrollo: lecciones de estudios comparados sobre insectos holometabolosos” Andrew Peel, 2008 en Transacciones filosóficas de la Royal Society, B. Publicado bajo The Royal Society Academic Institution Licencia de sitio único DOI: 10.1098/rstb.2007.2244 |
Genes de efecto materno: Bicoide, Nanos, Jorobado y un puñado de otros genes de Drosophila se conocen como genes de “efecto materno”. Esto significa que si los mutas en una mosca hembra, el fenotipo mutante se verá en su descendencia. Dado que ya sabes que los ARNm de Bicoid y Nanos son elaborados por las células de la madre y bombeados al oocito antes de la fertilización debería quedar bastante claro por qué es esto. Los genes de efecto materno configuran amplias regiones anterior y posterior en la mosca de la fruta así como regiones dorsal/ventrales (que no cubriremos aquí, aunque es una historia bastante jugosa).
Genes Gap: Los genes de efecto materno son factores de transcripción y traducción que afectan la expresión de los genes gap. Los genes gap son factores de transcripción que se expresan en tiras anchas a lo largo del eje A/P (Figura 2). Se llaman genes gap porque si uno es mutado conduce a una “brecha” en el embrión - a la larva le faltará una gran porción de su cuerpo 2.
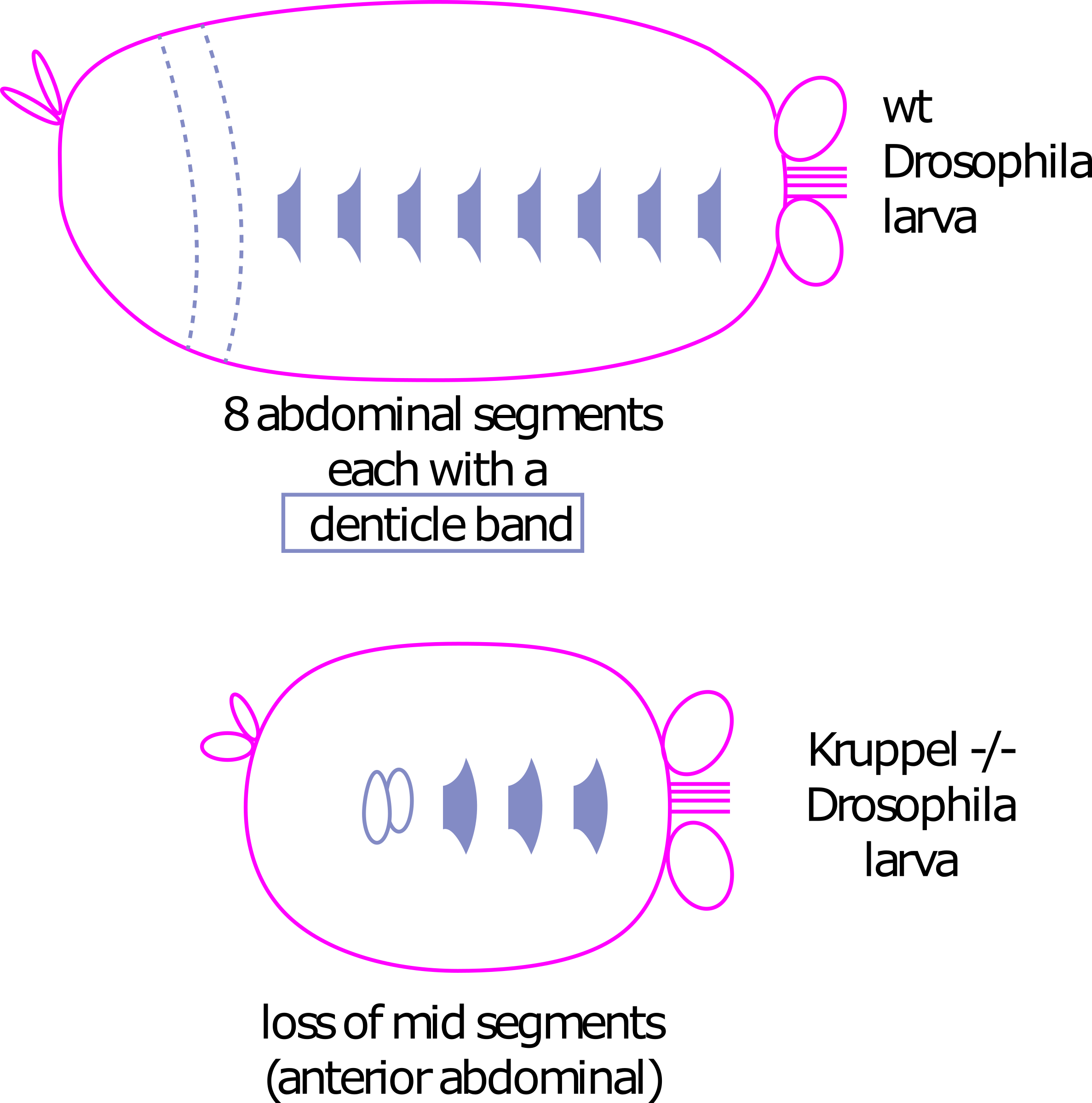 |
Figura 2: Genes de Gap. El panel izquierdo muestra la expresión de ARNm de tres genes de brecha de Drosophila, de Wikimedia commons por el usuario Celefin. Se trata de un derivado de una figura original de Haecker A, Qi D, Lilja T, Moussian B, Andrioli LP, Luschnig S, Mannervik, M. DOI 10.1371/journal.pbio.0050145 publicado bajo licencia CC BY 2.5 Panel derecho muestra el efecto de una mutación homocigótica de Kruppel en un embrión. Las formas grises son bandas dentulares (cerdas) que se encuentran en cada segmento abdominal. El mutante Kruppel ha perdido 6 de estas bandas, lo que corresponde con la expresión de Kruppel sobre los primeros 6 segmentos abdominales. Panel derecho de Amanda Lo y Ajna Rivera. Una versión editable svg de esta figura está disponible en https://scholarlycommons.pacific.edu/open-images/24/
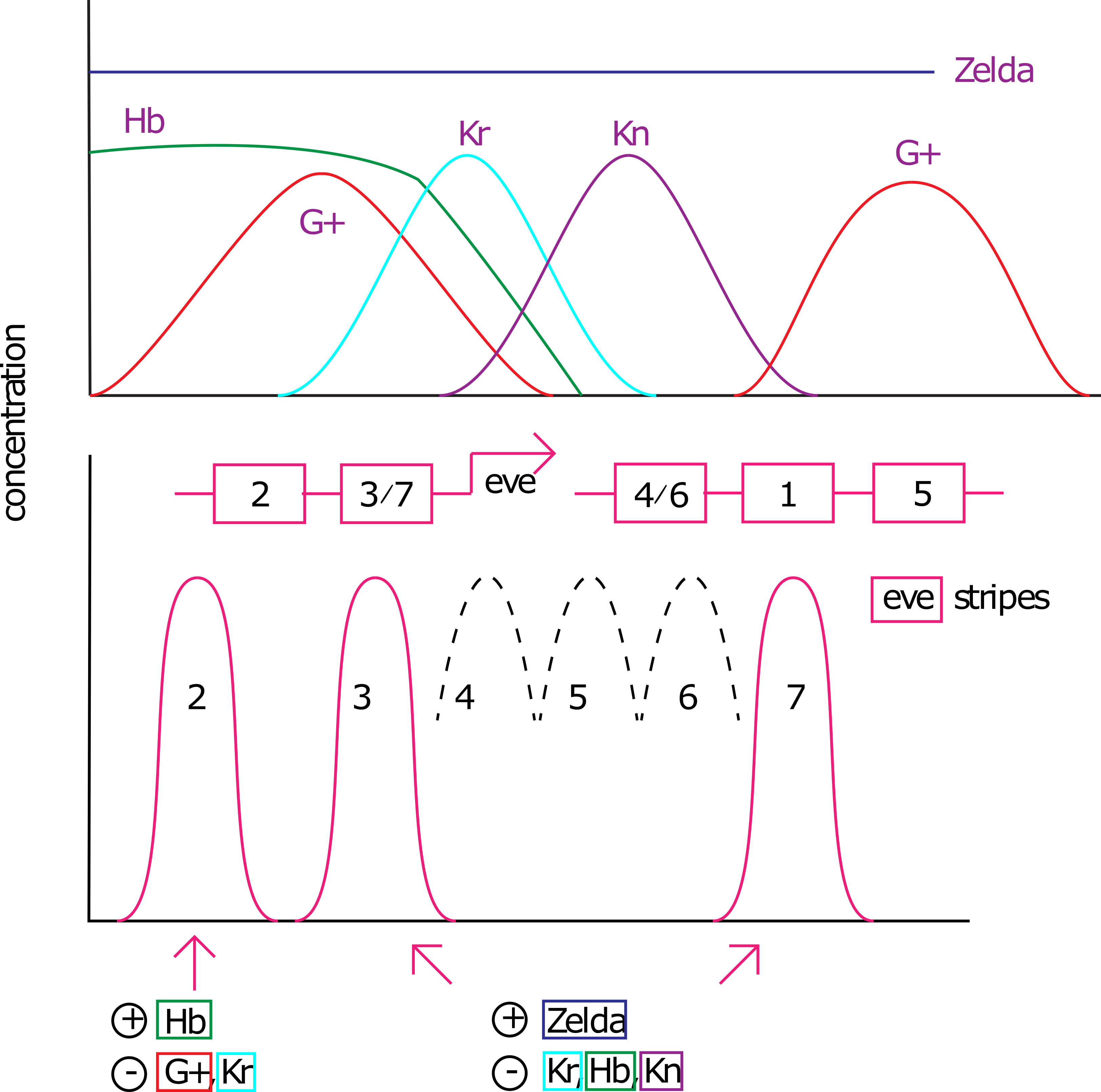
Genes de regla de pares: Los genes Gap y los genes de efecto materno afectan la expresión de los genes de regla de pares, los cuales se expresan en segmentos pares o impares 2. Estos son particularmente interesantes porque estos genes cierran la brecha entre la regionalización y la modularidad. Se expresan en un patrón modular, pero obtienen su aporte a partir de un patrón de regionalización. Genéticamente, esto significa que la expresión de cada uno de estos genes es activada por múltiples combinaciones de proteínas gap y de efecto materno. Por ejemplo, el gen saltado par se expresa en 7 franjas a lo largo del eje A/P. Cuenta con 4 principales potenciadores cis-reguladores que controlan esto. Un elemento que controla las bandas 3 y 7 tiene sitios de unión para Jorobado y Knirps, los cuales reprimen la expresión impulsada por el potenciador 3+7 (Figura 2). Este potenciador es activado por el activador ubicuo (expresado en todas partes) Zelda 3. El elemento que controla la banda 2 es aún más complejo con sitios de unión para Bicoid, Jorobado, Caudal, Kruppel y Knirps (Figura 3) 4.
Genes de polaridad de segmento: Los genes de polaridad de segmento se expresan en la parte anterior o posterior de cada segmento y ayudan a definir los límites del segmento así como el patrón A/P de cada segmento (Figura 4). Al igual que los genes de regla de pares, estos se expresan en franjas a lo largo de todo el eje A/P, más que en un patrón regional como los genes Maternal Effect y Gap. Los dominios de expresión de los genes de polaridad de segmento están regulados en gran medida por los genes de regla de pares y de brecha.
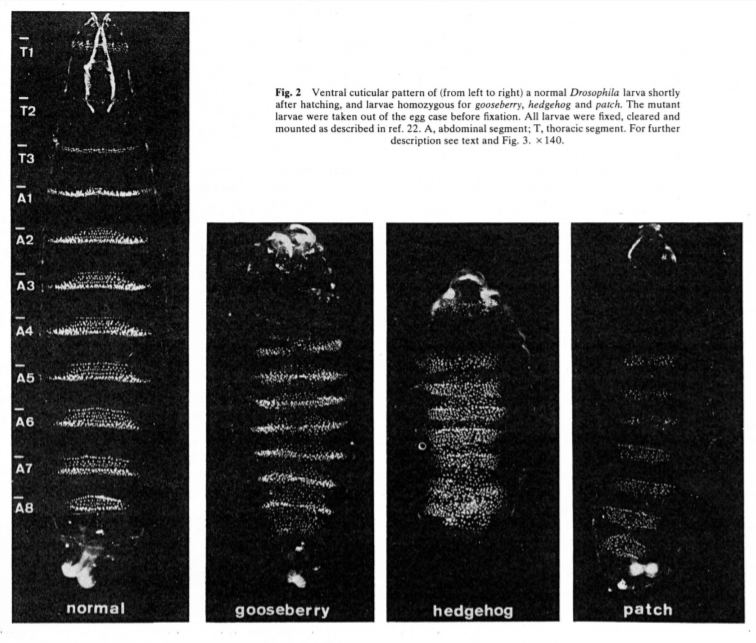
Genes Hox: Otro Nivel de Regionalización
Si bien los genes de efecto materno y los genes gap regionalizan en cierta medida el embrión de Drosophila, el animal adulto tendrá un cuerpo aún más regionalizado. El patrón del gen Hox proporciona regionalización a nivel de segmento. Especifica a dónde irán las piernas, alas, antenas, partes bucales, genitales y otras estructuras únicas (Figura 5). Como ya sabes, la mayoría de los animales usan los genes Hox para hacer patrones A/P y es un poco misterioso cómo y por qué tantas estrategias de desarrollo temprano diversas se fusionan en el mismo patrón A/P, y luego divergen en diversos planes corporales más adelante en el desarrollo.
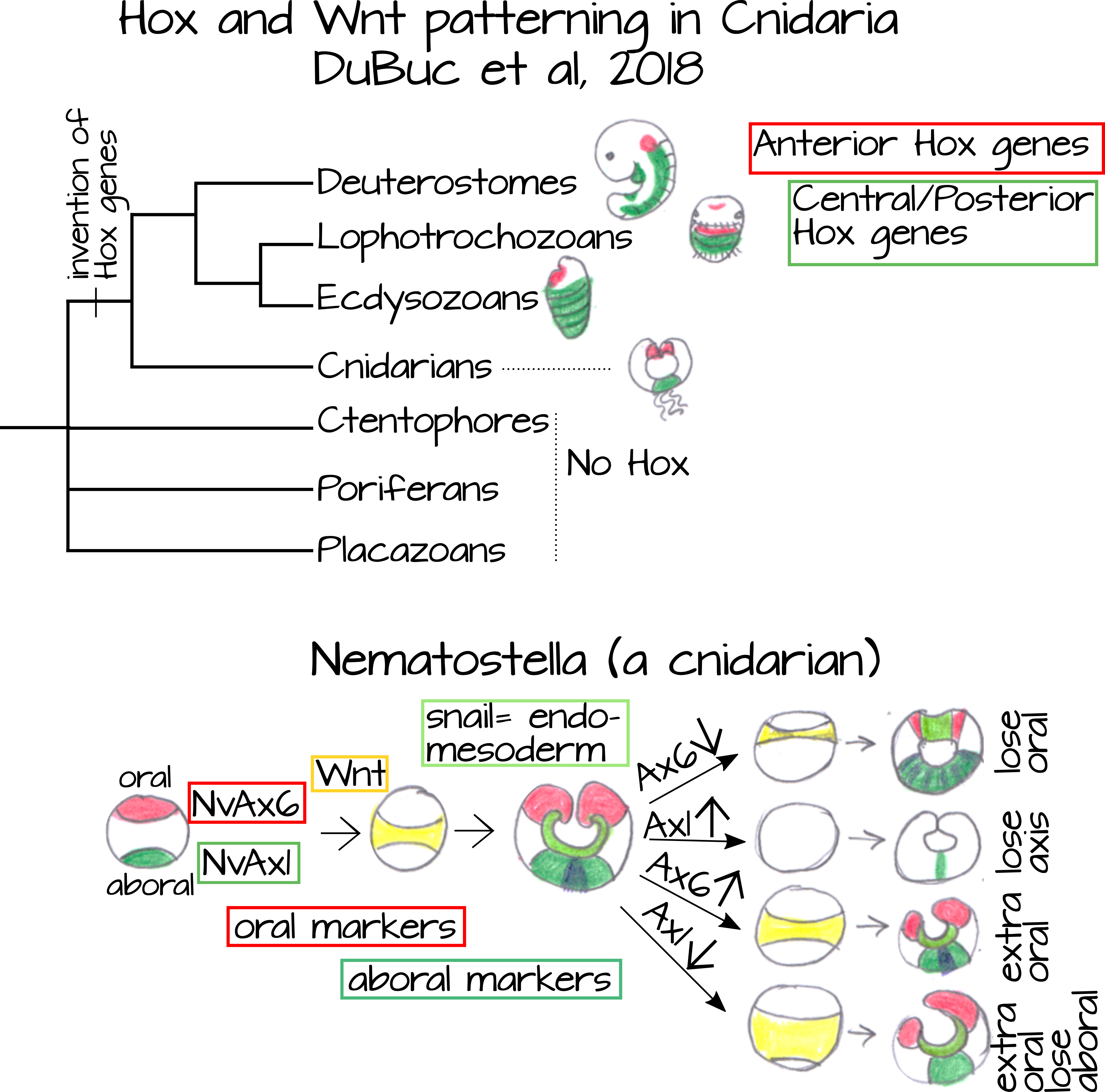
¿Qué tan conservado está el patrón Hox? Primero, casi todos los animales tienen genes Hox y muchos de ellos son sinténicos, lo que significa que el orden de los genes Hox se conserva a lo largo del cromosoma. Esto está relacionado con la regulación de los genes Hox, tienen factores reguladores distales (uno de los cuales veremos más adelante en Apéndices Ectodérmicos) pero también tienen factores reguladores proximales. En vertebrados, algunos genes en el cúmulo pueden compartir factores reguladores 5. Debido a esto, el orden genético importa. Una analogía es: si compartes un auto con tu vecino, no puedes mudarte a un nuevo barrio sin perder el acceso al auto. Este intercambio de elementos es una restricción evolutiva que ha dado como resultado la conservación de estos factores reguladores y la preservación de la síntesis de los genes Hox. En linajes invertebrados, la sintenia puede conservarse debido a la estrecha agrupación de los genes entre sí y sus elementos reguladores cis proximales 5. Sin embargo, algunas especies y linajes han perdido esta sintenía conservada, notablemente los nematodos y los tunicados 5. A pesar de estas pérdidas, la sintenía todavía parece ser la regla con al menos cierta conservación del orden genético que se encuentra en la mayoría de los filos examinados, incluyendo el filo que contiene HOX de mayor ramificación basal, los cnidarios 6.
Los cnidarios (como las jaleas y las anémonas de mar) son un caso interesante en el que observar los genes Hox porque si bien se sabe que los genes Hox son elementos de patrón A/P conservados, los cnidarios carecen de un eje A/P. ¿Cómo pueden faltar algo tan importante? Bueno es por cómo definimos anterior y posterior. Una definición estándar es que anterior está hacia la cabeza y posterior es hacia la cola. ¿Qué pasa con los animales que no tienen una cabeza obvia? Contamos su boca y cualquier conglomeración de órganos sensoriales principales como el extremo de la cabeza y seguimos moviéndose a lo largo de su eje largo hasta la punta más cercana del cuerpo. Típicamente, a lo largo del eje largo del cuerpo, la punta más cercana a la boca es el extremo anterior y la punta más cercana al ano es el extremo posterior. Los cnidarios, sin embargo, no sólo carecen de cabeza sino que también carecen de órganos sensoriales mayores y una boca. En cambio tienen un anillo de tentáculos (que pueden apuntar hacia arriba o hacia abajo dependiendo de la especie) que rodea una sola abertura que sirve de boca y ano.
Se han propuesto muchos escenarios evolutivos para homologizar el eje “radial” cnidario con los ejes bilaterianos A/P y D/V. Los genes Hox son una forma de ver esto. DuBuc et al 6 encontraron que los homólogas cnidarias de los genes Hox anteriores (rojo en las Figuras 6 y 7) se expresan en el extremo oral de un embrión de anémona (Cnidaria) y los homólogas cnidarios de genes Hox centrales/posteriores (azul/verde en la Figura 6) se expresan en el extremo aboral (el extremo opuesto a la boca/ano). También encontraron que Wnt, un gen de transducción de señales involucrado en muchos procesos de desarrollo, incluida la especificación anteroposterior, se expresó entre los dominios de expresión de Ax6, un gen Hox anterior, y Ax1, un gen Hox posterior