
10.1: Estudio de caso - Apéndices ectodérmicos
- Page ID
- 52856

Los apéndices ectodérmicos son órganos asociados a la piel como plumas y cabello. Por lo general, solo llamamos a las elaboraciones de piel de vertebrados “apéndices ectodérmicos” a pesar de que otros grupos de animales también tienen órganos de la piel (como glándulas). En esta sección solo nos centraremos en los apéndices vertebrados con un enfoque especial en las glándulas mamarias. Los detalles sobre esto así como un resumen del desarrollo de otros órganos de la piel se pueden encontrar aquí: Mecanismos de organogénesis ectodérmica.
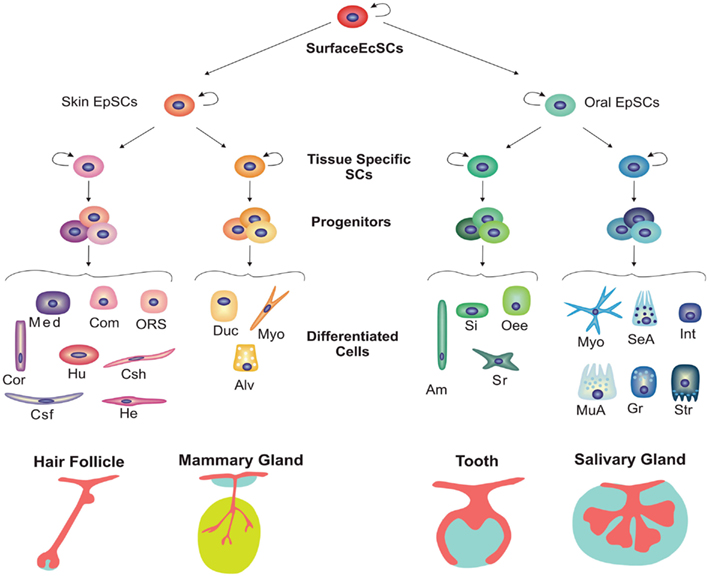
Los apéndices ectodérmicos (u órganos) tienen patrones de desarrollo comunes y se cree que también tienen un órgano evolutivo común, con la coopción de elementos de diferentes tipos de apéndices que ocurren con frecuencia. Estos apéndices son primero obvios como un engrosamiento de la epidermis para formar un “placode”. A esto le sigue la proliferación celular de la epidermis y la condensación de las células mesenquimales subyacentes de la dermis para formar un brote con una base mesenquimatosa gruesa. El brote derivado ectodérmico ahora puede experimentar una de múltiples trayectorias, que elige en base a señales del mesodermo, el mesénquima de la dermis así como el mesodermo somítico (Figura 1). Clasificamos aproximadamente estos apéndices en dos grupos principales: los apéndices orales, incluidos los dientes y las glándulas salivales, y los apéndices de la piel, incluyendo plumas, escamas, pelo, glándulas mamarias, glándulas sudoríparas y glándulas sebáceas.
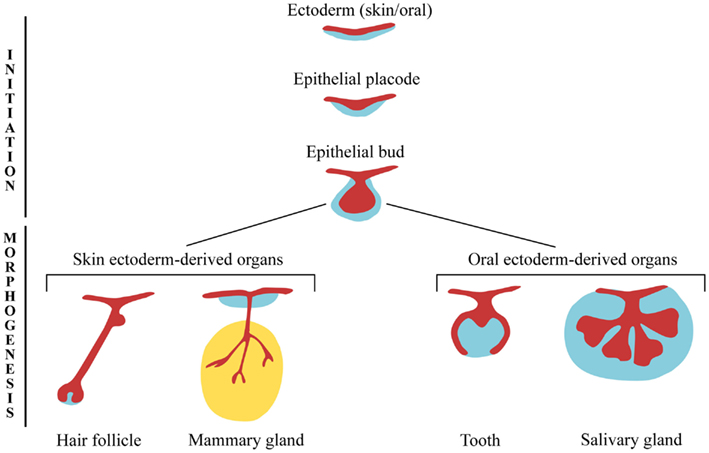
Al principio, los científicos notaron algo interesante: una vez que un placode ectodérmico estaba en su lugar, se podía inducir a formar uno de los muchos tejidos. Por ejemplo, el mesénquima del brote mamario puede estimular la formación de glándulas mamarias en el epitelio dorsal 2. Un experimento interesante mostró que el mesénquima dental de ratón podría incluso inducir dientes en el epitelio oral de pollo, a pesar de que las aves normalmente no crecen dientes 3. Como se discutió anteriormente, el desarrollo del apéndice ectodérmico sigue la trayectoria de general a específico (Figura 2) siendo los placodes tempranos menos especificados que los brotes posteriores.
Evolución y diversidad de apéndices cutáneos de tetrápodos
Los peces tienen sus propios apéndices cutáneos asombrosos y diversos, incluidos los órganos de “línea lateral” que detectan campos eléctricos, cambios de presión y flujo de agua 4. Sin embargo, aquí nos centraremos en los apéndices ectodérmicos de los tetrápodos con un enfoque en las glándulas mamarias de Mammalia. Se plantea la hipótesis de que el tetrápodo ancestral ha tenido glándulas mucosas en su piel, similares a las de los anfibios vivos. Estas glándulas mucosas mantienen la piel húmeda y evitan que las partes internas del animal se sequen. Adicionalmente, las glándulas mucosas y otras glándulas cutáneas son utilizadas por la cría de anfibios modernos para evitar que sus huevos sin cáscara de huevo se sequen en los ecosistemas terrestres y pueden haber jugado un papel similar en los tetrápodos tempranos (Figura 3).

Hace unos 350 millones de años, un grupo de tetrápodos, el Amniota, desarrolló un nuevo tipo de proteína de la piel que impedía que su piel se secara: la queratina. Todos los amniotes (incluyendo mamíferos, aves, lagartos, etc) tienen alfa-queratina y los saurópsidos (aves y otros reptiles) tienen además beta-queratina, que utilizan para construir plumas y escamas. A diferencia de los mamíferos ponedoras, las saurópsidas tienen cáscaras de huevo calcificadas que reducen la pérdida de agua. Los mamíferos ponedores de huevos (los monotremos como ornitorrinco), tienen huevos descascarados de pergamino que pueden secarse rápidamente. Se cree que esta es la condición ancestral para el grupo que contiene los mamíferos, los sinápsidos y las secreciones glandulares de la piel del progenitor melancólico (como las secreciones de moco en las salamandras) han ayudado a prevenir la desecación de huevos en sinápsidos ancestrales, incluidos los mamíferos 5.
El mamífero ancestral heredó así una piel glandular que era multifuncional: probablemente secretaba aceite, sudor y otras secreciones diluidas (posiblemente una forma de sudor) para mantener sus huevos hidratados. Podemos obtener algunas pistas sobre cómo estas glándulas cutáneas se diversificaron en múltiples apéndices ectodérmicos comparando los tres clados vivos de mammales, los monotremos ponedores de huevos, los marsupiales therianos y los placentarios therianos (también conocidos como los euterianos). Los mamíferos tienen cuatro tipos principales de glándulas cutáneas, estas son las glándulas productoras de cera del oído (que ignoraremos por ahora, las glándulas sebáceas que producen aceite, las glándulas sudoríparas y las glándulas mamarias. Las tres últimas de estas glándulas tienen relaciones anatómicas especiales entre sí. Un folículo piloso típico está asociado con una glándula sebácea y también puede asociarse con una glándula sudoríparas apocrina. Las glándulas mamarias en algunas especies están asociadas a un folículo piloso así como a una glándula sebácea. Esto ha llevado a la hipótesis de que las glándulas mamarias se derivan al menos en parte de las glándulas sudoríparas apocrinas (Figura 4).
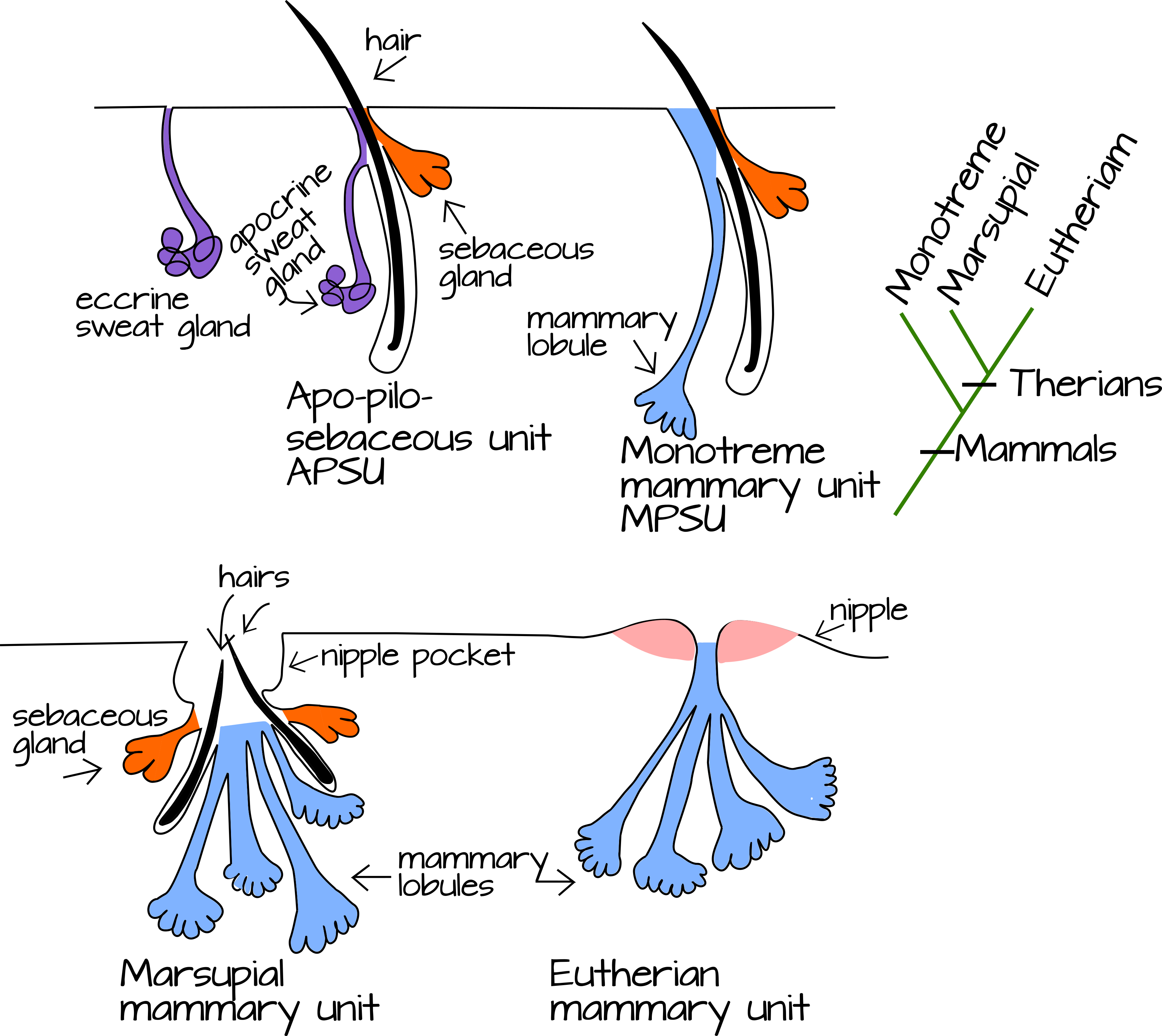 |
Figura 4: Comparación de glándulas cutáneas de mamíferos. La APSU (unidad apo-pilosebácea) es un tipo común de glándula cutánea que se encuentra en la axila y la ingle de los humanos. Consiste en una glándula sebácea, un folículo piloso y una glándula sudoríparas apocrina. Las glándulas apocrinas secretan al pellizcar parte de una célula secretora y liberarla en el conducto. Esto es muy diferente de las glándulas sudoríparas típicas (ecrinas) que secretan por exocitosis y se encuentran en toda la piel de los humanos. Se cree que la MPSU (unidad mammolobular-pilosebácea) es una APSU derivada con células secretoras mammolobulares apocrinas que reemplazan a las células de la glándula sudorosa. Una glándula mamaria contiene una o más MPSU y un pezón (en la mayoría de los casos, aunque algunos mamíferos como los monotremes carecen de pezones). |
Descripción general del desarrollo de la glándula mamaria
A pesar de la diversidad en sus formas finales, las glándulas mamarias se desarrollan a lo largo de vías en gran medida similares a través Como se muestra en la Figura 1, son inducidos por el mesénquima subyacente y otras señales para formar dos líneas mamarias paralelas, que probablemente son inducidas por la secreción de FGF10 inducida por GLI3 por un subconjunto del mesodermo somítico, impulsando la expresión de Wnt10b en el ectodermo suprayacente y especificando el destino de las células mamarias 6. Los monotremos retienen líneas mamarias como parches de células secretoras de leche. La diafonía entre la epidermis y la dermis (mesenquimatosa) genera un brote mamario, que crece hacia abajo en el mesénquima para formar un brote mamario. En los therianos, los brotes y brotes mamarios reciben señalización BMP4 del mesénquima subyacente para continuar impidiendo que tomen el destino predeterminado del folículo piloso 6. En otras partes de la piel, Noggin y Shh inhiben BMP4 y permiten el destino predeterminado del cabello 6. Los brotes mamarios epiteliales expresan PTHrP (Péptido Relacionado con la Hormona Paratiroidea), que activa su receptor en el mesénquima para inducir la condensación y diferenciación del brote primario mamario en crecimiento. Por lo tanto, existe una diafonía entre la epidermis y la dermis (mesénquima) para generar el brote mamario e inducir el crecimiento en el brote primario. En algunos mamíferos placentarios como los ratones, el brote primario crece profundamente en el tejido adiposo hipodérmico donde comienza a ramificarse en tejido glandular alveolar. En otros mamíferos se ramifica más superficialmente en brotes secundarios, que a su vez se ramifican en tejido glandular alveolar 8.
¿A dónde fue el pelo?
Si las glándulas mamarias evolucionaron a partir de los folículos pilosos y sus glándulas asociadas, ¿qué pasó con el cabello? Si bien muchos mamíferos placentarios tienen pelo aereolar (Figura 5), la mayoría no tiene pelo asociado con la parte del pezón que secreta leche. Además, ningún mamífero placentario examinado desarrolla un folículo piloso durante o antes de la formación de un brote mamario. Volveré al tema de los mamíferos placentarios en un momento, pero primero me gustaría pasar un poco de tiempo mirando a nuestros primos peludos aerola.
Los monotremos, los mamíferos basalmente ramificados, mantienen la asociación capilar con las glándulas mamarias (Figura 4). Oftedal propone que este es el caso ancestral de los mamíferos y que los pelos mamarios, más que un pezón, tienen un doble uso: no solo extraen la leche de las glándulas de la piel a la boca de un lactante, sino que también extraen secreciones húmedas sobre huevos propensos a la desecación. La evolución del desarrollo embrionario interno y el nacimiento vivo en los mamíferos therianos cambiaron la presión evolutiva sobre las glándulas productoras de leche. Perdieron la presión selectiva para mantener los huevos hidratados y en su lugar fueron seleccionados únicamente para la nutrición de los infantes. Esto pudo haber impulsado los conductos de leche a condensarse y fluir hacia un único pezón por glándula mamaria, en lugar de un parche de pelo disperso (Figura 6) 5,8.
La evidencia del desarrollo de la glándula mamaria en mamíferos therianos da cierto apoyo a esta hipótesis. En los marsupiales, el desarrollo temprano de la glándula mamaria procede como una versión físicamente condensada del desarrollo monotreme. En monotremes, un solo brote primario mamario se desarrolla en un folículo piloso, una glándula sebácea y células secretoras y ductales mamarias (la MPSU multicomponente). En los marsupiales, un solo brote primario forma múltiples MPSU, cada uno con su propio folículo piloso, glándula sebácea y células mamarias. Sin embargo, los folículos pilosos eventualmente se atrofian y mueren, dejando atrás un gran bolsillo en el que la leche y el sebo pueden vaciar (Figura 4) 8. Los mamíferos placentarios (euterianos), como nosotros, son más variables. Los euterianos pueden producir uno o muchos brotes primarios por bulbo mamario y (como se mencionó anteriormente) pueden producir o carecer de brotes secundarios. Las especies sin brotes secundarios no tienen folículos pilosos y sebáceas asociados con el crecimiento de las glándulas mamarias, pero las especies con brotes secundarios sí. Esto incluye humanos y caballos. Nuestros folículos pilosos y glándulas sebáceas MPSU se atrofian y mueren durante la embriogénesis por lo que nos quedamos con una MPSU hecha de un solo componente: los alvéolos y conductos mamarios 8.
Al comparar el desarrollo de las glándulas mamarias de estos diferentes grupos, Oftedal y Dhouailly han propuesto un escenario para la evolución de las glándulas mamarias por coopción y heterocronía. Brevemente, las glándulas mamarias utilizan mecanismos de desarrollo de placode cutáneos existentes para comenzar a formarse, en particular el patrón existente utilizado por las glándulas apo-pilosebáceas, luego cambian a un programa de morfogénesis de ramificación (un mecanismo de patrón del desarrollo que se encuentra en el riñón, los pulmones y las glándulas salivales en tetrápodos pero también en plumas de aves y desarrollo traqueal de insectos). Esto crea el árbol ductal de la glándula mamaria como la porción apocrina de la unidad apo-pilosebácea, en lugar de una glándula sudoríparas 8. El origen del pezón suprayacente aún no está claro, pero es inducido por el desarrollo de la glándula mamaria.
La genética de la evolución de la glándula mamaria: coopción y novedad
Anteriormente, discutí brevemente la elección del destino que hace la piel entre el pelo y la glándula mamaria: activar la señalización BMP4 en las células mesenquimales a través de la señalización FGF10 de parte del mesodermo somítico apaga el programa capilar predeterminado y enciende el programa mamario. Las interacciones FGF/Wnt/BMP se utilizan en la formación de otra piel en ectodermo suprayacente, se usa para modelar otros apéndices cutáneos y puede ser un casete general para el patrón ectodérmico 9. De esta manera, las glándulas mamarias placode y la formación de yemas probablemente son impulsadas por una señalización similar a la de otros apéndices ectodérmicos con algunas modificaciones. Una vez que el placode/brote mamario comienza a crecer, expresa un gen común del órgano epidérmico: PTHrP. La PTHrP tiene una función más ancestral en vertebrados en formación ósea endocondrial y erupción dental. Como los dientes son una estructura epitelial ectodérmica, es probable que el papel de PTHrP en el desarrollo de la glándula mamaria se coopte del desarrollo dental. La evidencia de coopción de genes dentales proviene de otra familia de genes clave en el desarrollo dental: la proteína de unión al calcio ODAM. La ODAM se duplicó durante la evolución del tetrápodo para formar las caseínas, las proteínas transportadoras de calcio en la leche 10
FGF10 y Wnt10b no son los únicos jugadores en especificación mamaria. Varios genes Hox están implicados en la especificación de órganos ectodérmicos, con Hoxc8 iniciando el desarrollo de placode mamario de ratón y Hoxb3, b6, b9, d9, d10 y d8, entre otros, expresados durante el desarrollo posterior, incluso durante el desarrollo mamario posnatal 11,12. De estos, Hoxd9 y d8 se expresan con mayor frecuencia en bulbos mamarios de ratón. Estos dos genes también están involucrados en el desarrollo tanto de las extremidades como del cabello en ratones.
Mientras que Hoxd9 y d8 se localizan uno al lado del otro en el cromosoma y en gran medida regulados por dos elementos cis-reguladores gigantes Hoxd-T-Dom y HoxD-C-Dom, no exhiben patrones de expresión completamente idénticos, lo que sugiere que existen reguladores específicos que actúan diferencialmente en su individuo promotores. Recientemente, científicos del laboratorio Beccari en Ginebra diseccionaron los elementos reguladores que impulsan la expresión de Hox d8 y d9 y encontraron cómo lograron pasar de expresarse en la extremidad y el pelo a expresarse en el bulbo mamario. Mientras que la expresión del bulbo mamario Hoxd8 dependía de una región potenciadora local de 13kb, la expresión del bulbo mamario Hoxd9 dependió de regiones reguladoras cis distantes en T-DOM 12.
El elemento regulador cis del bulbo mamario Hoxd9 es particularmente intrigante porque parece depender en parte de los elementos reguladores cis existentes que impulsan la expresión de Hoxd9 en las extremidades. La expresión de las extremidades es impulsada por elementos reguladores cis dentro del T-DOM y durante el desarrollo de la extremidad, una región gigante de 1500 kb del cromosoma se pliega para contactar con Hoxd9. El potenciador de yemas mamarias para Hoxd9 también se encuentra dentro de esta región de 1500 kb, y se piensa que este potenciador aprovecha la atracción entre el potenciador de yemas de las extremidades y el cúmulo de Hox para acceder a Hoxd9 durante el desarrollo del bulbo mamario (Figura 7) 12. Tomados en conjunto, esto comienza a pintar una imagen de la evolución de los placodes mamarios y bulbos: utilizan el factor de transcripción existente y los genes de patrones en nuevas combinaciones. Al menos uno de los genes involucrados se expresa en el tejido mamario en desarrollo mediante la cooptación y modificación de elementos potenciadores existentes.
Una vez construidas las glándulas mamarias embrionarias, todavía necesitan secretar leche. Si bien las similitudes entre las glándulas productoras de leche y las glándulas sudoríparas apocrinas son obvias a nivel anatómico y de desarrollo, el contenido de las dos sustancias producidas es sustancialmente diferente. Un producto lácteo, las caseínas, está hecho de proteína dental ODAM mutada (ver arriba). La principal proteína esencial en el procesamiento de las gotitas de grasa que se encuentran en la leche, XOR, suele ser una proteína antimicrobiana pero juega un segundo papel en la biosíntesis de la leche. Este es un caso claro de heterotopía (expresión de un gen en un nuevo dominio). La lisozima tomó ambos caminos: se cooptó del sistema inmune como una proteína antimicrobiana en las glándulas mamarias, y también se sometió a una duplicación génica para formar una proteína de suero utilizada para la lactosa 13. Aunque la composición de la leche es compleja y varía de una especie a otra (incluso dentro de una especie a medida que un bebé envejece), podemos rastrear la historia evolutiva de estos componentes y ver cómo la evolución podría haber actuado para producir esta secreción.
Referencias
- “Determinación del destino de células madre durante el desarrollo y regeneración de órganos ectodérmicos”. (2012) Fronteras en Fisiología, Lucía Jiménez-Rojo, Zoraide Granchi, Daniel Graf y Thimios A. Mitsiadis, https://doi.org/10.3389/fphys.2012.00107.
- “Expresión fenotípica mamaria inducida en células epidérmicas por mesénquima mamaria embrionaria” (1995) Gerald R. Cunha, Peter Young, Konstantin T Christov, R C Guzmán, S. B. Nandi, Frank J. Talamantes, Gudmundur Thordarson, PMID: 7572029.
- “Desarrollo de dientes en embriones de pollo después de trasplantes de cresta neural de ratón”, (2003), PNAS, Thimios A. Mitsiadis, Yvonnick Chéraud, Paul Sharpe, y Josiane Fontaine-Pérus, https://doi.org/10.1073/pnas.1137104100.
- “La línea lateral puede mediar la reotaxis en peces” (1997), Nature, John C. Montgomery, Cindy F. Baker, y Alexander G. Carton, https://doi.org/10.1038/40135.
- “El origen de la lactancia como fuente de agua para huevos sin cáscara de pergamino” (2002) Journal of Mammary Gland Biology and Neoplasia, Olav T. Oftedal, https://doi.org/10.1023/A:1022848632125. .
- “Un nuevo escenario para el origen evolutivo del cabello, la pluma y las escamas aviares”, (2009) Journal of Anatomy, Danielle Dhouailly, doi: 10.1111/j.1469-7580.2008.01041.x.
- “La proteína relacionada con la hormona paratiroidea mantiene el destino epitelial mamario y desencadena la diferenciación de la piel del pezón durante el desarrollo mamario embrionario.” (2001) Development, J. Foley, P. Dann, J. Hong, J. Cosgrove, B. Dreyer, D. Rimm, M. Dunbar, W. Philbrick, J. Wysolmerski, PMID: 11171335.
- “Evo-devo de la glándula mamaria”, (2013), Revista de Biología y Neoplasia de Glándula Mamaria, Olav T. Oftedal y Danielle Dhouailly, doi: 10.1007/s10911-013-9290-8.
- “Se requiere señalización FGF para iniciar el desarrollo de placode pluma” (2004) Development, Markus Mandler, Annette Neubüser, doi: 10.1242/dev.01203.
- “La evolución de los genes de caseína láctea a partir de genes dentales antes del origen de los mamíferos” (2012), Kazuhiko Kawasaki, Anne-Gaelle Lafont, Jean-Yves Sire, Molecular Biology and Evolution, https://doi.org/10.1093/molbev/msr020.
- “HOXC8 inicia un programa mamario ectópico regulando la expresión de FGF10 y Tbx3, y señalización Wnt/β-catenina” (2015) Desarrollo, Lara S. Carroll, Mario R. Capecchi, doi: 10.1242/dev.128298.
- “Control de la transcripción del gen Hoxd en el capullo mamario secuestrando un panorama regulatorio preexistente”, (2016), PNAS, Ruben Schep, Anamaria Necsulea, Eddie Rodríguez-Carballo, Isabel Guerreiro, Guillaume Andrey, Thi Hanh Nguyen Huynh, Virginie Marcet, Jozsef Zákány, Denis Duboule y Leonardo Beccari, https://doi.org/10.1073/pnas.1617141113.
- “¿Evolución de la glándula mamaria a partir del sistema inmune innato?” (2006) Bioensayos, Claudia Vorbach Mario R. Capecchi Josef M. Penninger, doi.org/10.1002/bies.20423.
Miniaturas: Imagen de primer plano de plumas de pavo real, que es un ejemplo de Apéndices Ectodérmicos. (Dominio Público).