
10.2: El modelo de nacimiento-muerte
- Page ID
- 53887

Un modelo de nacimiento-muerte es un proceso continuo de Markov que a menudo se utiliza para estudiar cómo el número de individuos en una población cambia a través del tiempo. Para la macroevolución, estos “individuos” suelen ser especies, a veces llamadas “linajes” en la literatura. En un modelo de nacimiento-muerte, pueden ocurrir dos cosas: nacimientos, donde el número de individuos aumenta en uno; y muertes, donde el número de individuos disminuye en uno. Suponemos que no más de un nuevo individuo puede formarse (o morir) durante un evento cualquiera. En términos filogenéticos, eso significa que los árboles de nacimiento-muerte no pueden tener “politomías duras”; cada evento de especiación da como resultado exactamente dos especies descendientes.
En la macroevolución, aplicamos el modelo de nacimiento-muerte a las especies, y típicamente consideramos un modelo donde cada especie tiene una probabilidad constante de dar a luz (especiar) o morir (extinguirse). Denotamos la tasa de natalidad por linaje como λ y la tasa de mortalidad por linaje como μ. Por ahora consideramos que estas tarifas son constantes, pero vamos a relajar esa suposición más adelante en el libro.
Podemos entender el comportamiento de los modelos de nacimiento-muerte si consideramos el tiempo de espera entre sucesivos eventos de especiación y extinción en el árbol. Imagínese que estamos considerando un solo linaje que existe en el tiempo t 0. Podemos pensar en el tiempo de espera para el siguiente evento, que será un evento de especiación que divide ese linaje en dos (Figura 10.2A) o un evento de extinción que marcará el final de ese linaje (Figura 10.2B). Bajo un modelo de nacimiento-muerte, ambos eventos siguen un proceso de Poisson, de manera que el tiempo de espera esperado para un evento sigue una distribución exponencial (Figura 10.2C). El tiempo de espera esperado para el siguiente evento de especiación es exponencial con el parámetro λ, y el tiempo de espera esperado hasta el siguiente evento de extinción exponencial con el parámetro μ. [Por supuesto, solo uno de estos puede ser el próximo evento. El tiempo de espera esperado para el siguiente evento (de cualquier tipo) es exponencial con el parámetro λ + μ, y la probabilidad de que ese evento sea especiación es λ/(μ + λ), extinción μ/(μ + λ)].
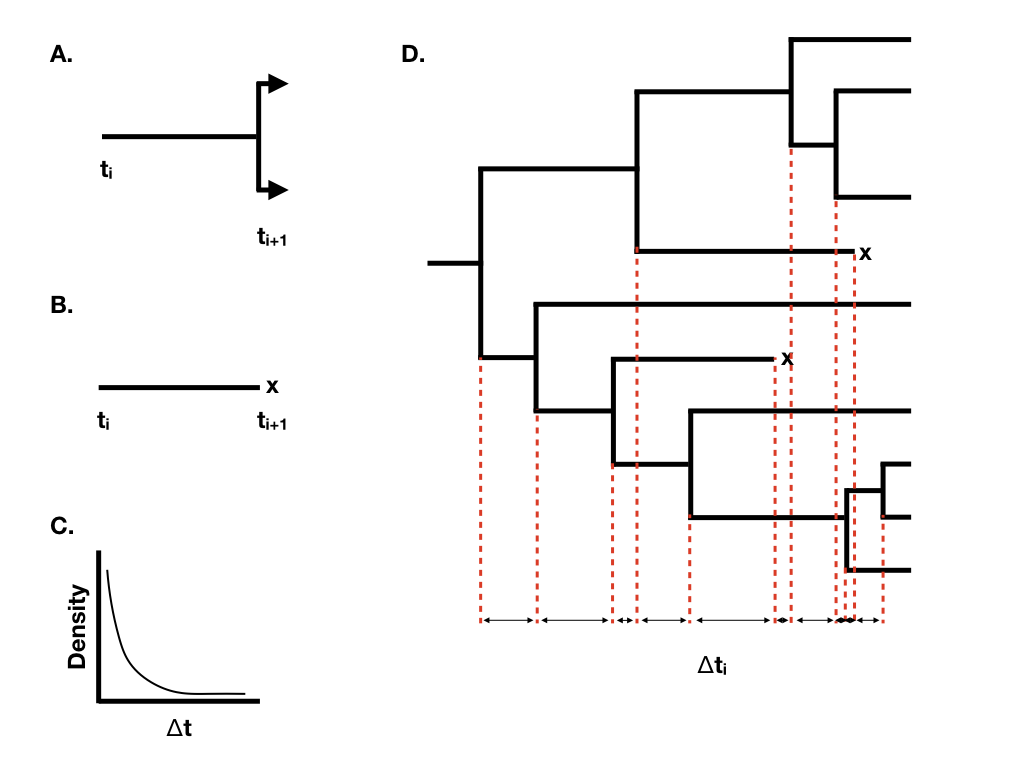
Cuando tenemos más de un linaje “vivo” en el árbol en cualquier momento, entonces cambia el tiempo de espera para el siguiente evento, aunque su distribución sigue siendo exponencial. En general, si hay N (t) linajes vivos en el tiempo t, entonces el tiempo de espera para el siguiente evento sigue una distribución exponencial con el parámetro N (t) (λ + μ), con la probabilidad de que ese evento sea especiación o extinción lo mismo que se ha dado anteriormente. Se puede ver a partir de esta ecuación que el parámetro de tasa de la distribución exponencial se hace mayor a medida que aumenta el número de linajes. Esto significa que los tiempos de espera esperados en todos los linajes se hacen cada vez más cortos a medida que se acumulan más linajes.
Mediante este enfoque, podemos cultivar árboles filogenéticos de cualquier tamaño (Figura 10.2D).
Podemos derivar algunas propiedades importantes del proceso de nacimiento-muerte en los árboles. Para ello, es útil definir dos parámetros adicionales, la tasa neta de diversificación (r) y la tasa de extinción relativa ():
\[r = λ − μ \label{10.A}\]
\[ \epsilon = \frac{\mu}{\lambda} \lable{10.1B}\]
Estos dos parámetros simplifican algunas de las ecuaciones siguientes, y también se encuentran comúnmente en la literatura.
Para derivar algunas propiedades generales del modelo de nacimiento-muerte, primero consideramos el proceso a lo largo de un pequeño intervalo de tiempo, Δ t. Suponemos que este intervalo es tan corto que contiene como máximo un solo evento, ya sea especiación o extinción (el intervalo también podría no contener eventos en absoluto). La probabilidad de especiación y extinción en el intervalo de tiempo se puede expresar como:
\[Pr_{speciation} = N(t)λΔt \label{10.2A}\]
\[Pr_{extinction} = N(t)μΔt \label{10.2B}\]
Ahora consideramos el número total de especies vivas en algún momento t, y escribimos esto como N (t). Es útil pensar en el valor esperado de N (t) bajo un modelo de nacimiento-muerte [consideramos la distribución completa de N (t) a continuación]. El valor esperado de N (t) después de un pequeño intervalo de tiempo Δ t es:
\[N(t + Δt)=N(t)+N(t)λΔt − N(t)μΔt \label{10.3}\]
Podemos convertir esto en una ecuación diferencial restando N (t) de ambos lados, luego dividiendo por Δ t y tomando el límite como Δ t se vuelve muy pequeño:
\[dN/dt = N(λ − μ) \label{10.4}\]
Podemos resolver esta ecuación diferencial si establecemos una condición de límite que N (0) = n 0; es decir, en el tiempo 0, comenzamos con n 0 linajes. Luego obtenemos:
\[N(t)=n_0e^{λ − μ}t = n_0e^{rt} \label{10.5}\]
Esta ecuación determinista nos da el valor esperado para el número de especies a través del tiempo bajo un modelo de nacimiento-muerte. Observe que el número de especies crece exponencialmente a través del tiempo siempre que λ > μ, es decir r > 0, y decae de otra manera (Figura 10.3).
También nos interesa el comportamiento estocástico del modelo, es decir, ¿cuánto debemos esperar que N (t) varíe de una réplica a otra? Podemos calcular la distribución de probabilidad completa para N (t), que escribimos como p n (t) = P r [N (t) = n] para todos n ≥ 0, para describir completamente el Comportamiento del modelo de nacimiento-muerte. Para derivar esta distribución de probabilidad, podemos comenzar con un conjunto de ecuaciones, una por cada valor de n, para hacer un seguimiento de las probabilidades de n linajes vivos en el tiempo t. Denotaremos cada uno de estos como p n (t) (hay un conjunto infinito de tales ecuaciones, de p 0 a p ∞). Entonces podemos escribir un conjunto de ecuaciones de diferencia que describen las diferentes formas en que uno puede alcanzar cualquier estado en un pequeño intervalo de tiempo Δ t. De nuevo asumimos que Δ t es suficientemente pequeño para que pueda ocurrir como máximo un evento (un nacimiento o una muerte). Como ejemplo, considere lo que puede suceder para hacer n = 0 al final de un cierto intervalo de tiempo. Hay dos posibilidades: o ya estábamos en n = 0 al inicio del intervalo de tiempo y (por definición) no pasó nada, o estábamos en n = 1 y el último linaje sobreviviente se extinguió. Escribimos esto como:
(eq. 10.6)
p 0 (t + Δ t) = p 1 (t) μ Δ t + p 0 (t)
De igual manera, podemos llegar a n = 1 ya sea comenzando con n = 1 y sin eventos, o pasando de n = 2 vía extinción.
(eq. 10.7)
p 1 (t + Δ t) = p 1 (t) (1 − (λ + μ)) Δ t + p 2 (t) 2 μ Δ t
Finalmente, cualquiera para n > 1, podemos alcanzar el estado de n linajes de tres maneras: desde un nacimiento (de n − 1 a n), una muerte (de n + 1 a n), o ninguna (de n a n).
(eq. 10.8)
p n (t + Δ t) = p n − 1 (t) (n − 1) λ Δ t + p n + 1 (t) ( n + 1) μ Δ t + p n (t) (1 − n (λ + μ)) Δ t
Podemos convertir este conjunto de ecuaciones de diferencia en ecuaciones diferenciales restando p n (t) de ambos lados, luego dividiendo por Δ t y tomando el límite como Δ t se vuelve muy pequeño. Entonces, cuando n = 0, usamos 10.6 para obtener:
(eq. 10.9)
d p 0 (t)/d t = μ p 1 (t)
Desde 10.7:
(eq. 10.10)
d p 1 (t)/d t = 2 μ p 2 (t) − (λ − μ) p 1 (t)
y de 10.8, para todos n > 1:
(eq. 10.11)
d p n (t)/d t = (n − 1) λ p n − 1 (t) + (n + 1) μ p n + 1 ( t) − n (λ + μ) p n (t)
Entonces podemos resolver este conjunto de ecuaciones diferenciales para obtener la distribución de probabilidad de p n (t). Usando la misma condición de contorno, N (0) = n 0, tenemos p 0 (t) =1 para n = n 0 y 0 de lo contrario. Entonces, podemos encontrar la solución a las ecuaciones diferenciales 10.9, 10.10 y 10.11. La derivación de la solución a este conjunto de ecuaciones diferenciales está más allá del alcance de este libro (pero ver Kot 2001 para una buena explicación de las matemáticas). Una solución fue obtenida por primera vez por Bailey (1964), pero usaré la forma equivalente más simple de Foote et al. (1999). Para p 0 (t) —es decir, la probabilidad de que todo el linaje se haya extinguido en el tiempo t— tenemos:
(eq. 10.12)
p 0 (t) = α 0 n
Y para todos n ≥ 1:
(eq. 10.13)
\[ p_n(t) = \sum\limits_{j=1}^{min(n_0,n)} \binom{n_0}{j} \binom{n-1}{j-1} \alpha^{n_0 - j} \beta^{n-j} [(1-\alpha)(1-\beta)]^j \]
Donde α y β se definen como:
(eq. 10.14)
\[ \alpha=\frac{\epsilon (e^rt-1)}{(e^rt-\epsilon)} \]
\[ \beta =\frac{(e^rt-1)}{(e^rt-\epsilon)} \]
α es la probabilidad de que algún linaje en particular se haya extinguido antes del tiempo t.
Tenga en cuenta que cuando n 0 = 1 —es decir, cuando empezamos con un solo linaje- las ecuaciones 10.12 y 10.13 simplifican a (Raup 1985):
(eq. 10.15)
p 0 (t) = α
Y para todos n ≥ 1:
(eq. 10.16)
p n (t) =( 1 − α) (1 − β) β i − 1
En todos los casos el número esperado de linajes en el árbol es exactamente como se indicó anteriormente en la ecuación (10.5), pero ahora tenemos la distribución de probabilidad completa del número de linajes dados n 0, t, λ y μ. Algunas parcelas capturan la forma general de esta distribución (Figura 10.4).
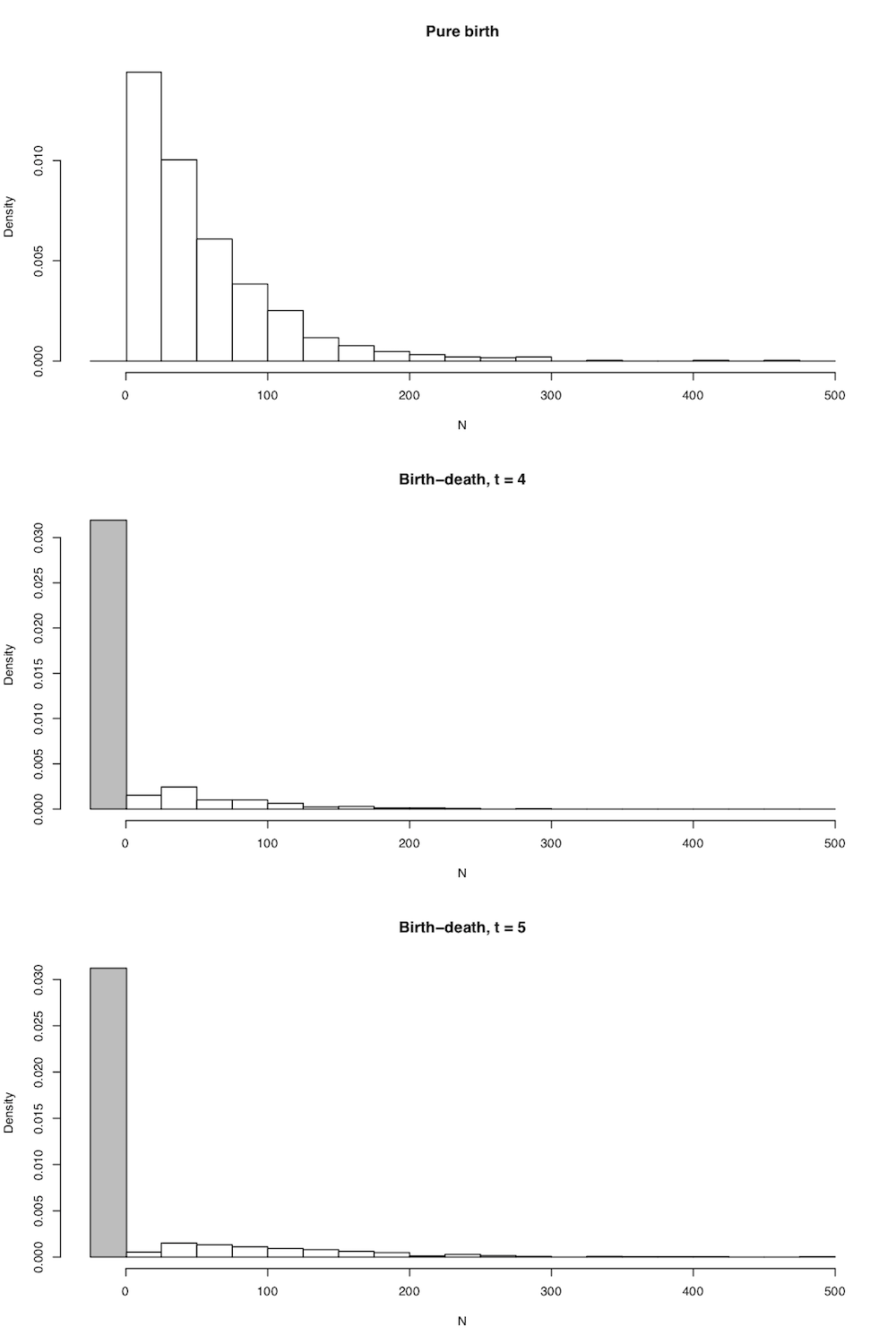
Existen bastantes métodos comparativos que utilizan la riqueza y la edad de especies de clados junto con las distribuciones definidas en 10.14 y 10.15 para hacer inferencias sobre las tasas de diversificación de clados (ver capítulo 11).