
17.3: Inmunidad adaptativa
- Page ID
- 53498

La respuesta inmune adaptativa o adquirida toma días o incluso semanas para que se establezca, mucho más tiempo que la respuesta innata; sin embargo, la inmunidad adaptativa es más específica para un patógeno invasor. La inmunidad adaptativa es una inmunidad que ocurre después de la exposición a un antígeno ya sea de un patógeno o de una vacunación. Un antígeno es una molécula que estimula una respuesta en el sistema inmune. Esta parte del sistema inmune se activa cuando la respuesta inmune innata es insuficiente para controlar una infección. De hecho, sin información del sistema inmune innato, no se pudo movilizar la respuesta adaptativa. Existen dos tipos de respuestas adaptativas: la respuesta inmune mediada por células, que es controlada por células T activadas, y la respuesta inmune humoral, que es controlada por células B activadas y anticuerpos. Las células T y B activadas cuyos sitios de unión a la superficie son específicos de las moléculas en el patógeno aumentan enormemente en número y atacan al patógeno invasor. Su ataque puede matar patógenos directamente o pueden secretar anticuerpos que potencian la fagocitosis de patógenos e interrumpen la infección. La inmunidad adaptativa también implica una memoria para dar al huésped protección a largo plazo contra la reinfección con el mismo tipo de patógeno; al reexponer, esta memoria del huésped facilitará una respuesta rápida y poderosa.
Células B y T
Los linfocitos, que son glóbulos blancos, se forman con otros glóbulos sanguíneos en la médula ósea roja que se encuentran en muchos huesos planos, como el hombro o los huesos pélvicos. Los dos tipos de linfocitos de la respuesta inmune adaptativa son las células B y T (Figura\(\PageIndex{1}\)). Que un linfocito inmaduro se convierta en una célula B o una célula T depende de en qué parte del cuerpo madura. Las células B permanecen en la médula ósea para madurar (de ahí el nombre “B” para “médula ósea”), mientras que las células T migran al timo, donde maduran (de ahí el nombre “T” para “timo”).
La maduración de una célula B o T implica volverse inmunocompetente, lo que significa que puede reconocer, por unión, una molécula o antígeno específico (discutido más adelante). Durante el proceso de maduración, las células B y T que se unen demasiado fuertemente a las propias células del cuerpo se eliminan para minimizar una respuesta inmune contra los propios tejidos del cuerpo. Quedan aquellas células que reaccionan débilmente a las propias células del cuerpo, pero que tienen receptores altamente específicos en sus superficies celulares que les permiten reconocer una molécula extraña, o antígeno. Este proceso ocurre durante el desarrollo fetal y continúa a lo largo de la vida. La especificidad de este receptor está determinada por la genética del individuo y está presente antes de que una molécula extraña se introduzca en el cuerpo o se encuentre. Así, es la genética y no la experiencia lo que inicialmente proporciona una amplia gama de células, cada una capaz de unirse a una molécula extraña específica diferente. Una vez que son inmunocompetentes, las células T y B migrarán al bazo y ganglios linfáticos donde permanecerán hasta que sean llamadas durante una infección. Las células B están involucradas en la respuesta inmune humoral, que se dirige a patógenos sueltos en la sangre y la linfa, y las células T están involucradas en la respuesta inmune mediada por células, que se dirige a las células infectadas.

Respuesta inmunológica humoral
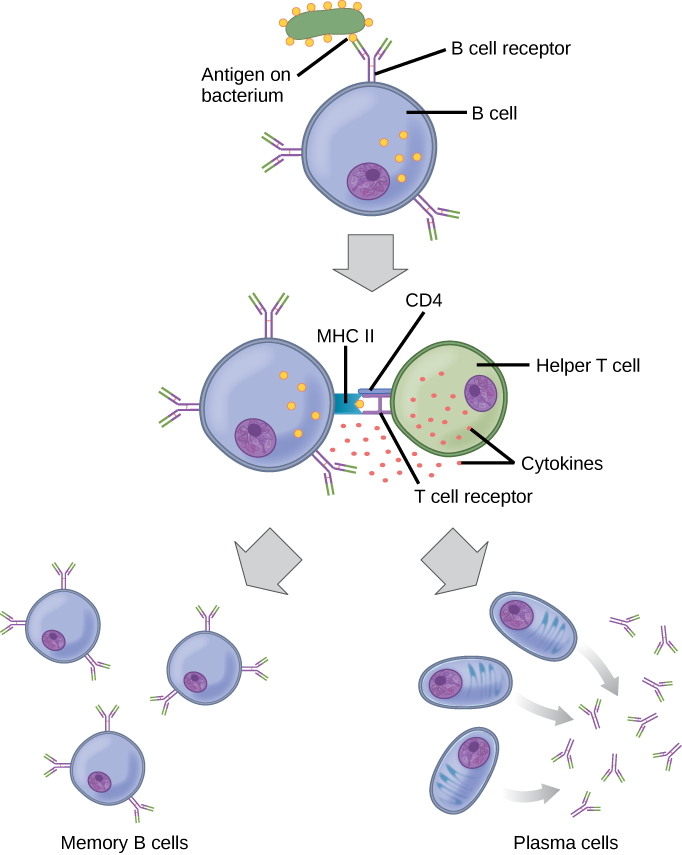
Como se mencionó, un antígeno es una molécula que estimula una respuesta en el sistema inmune. No todas las moléculas son antigénicas. Las células B participan en una respuesta química a los antígenos presentes en el cuerpo al producir anticuerpos específicos que circulan por todo el cuerpo y se unen con el antígeno cada vez que se encuentra. Esto se conoce como la respuesta inmune humoral. Como se discutió, durante la maduración de las células B, se produce un conjunto de células B altamente específicas que tienen muchas moléculas receptoras de antígeno en su membrana (Figura\(\PageIndex{2}\)).
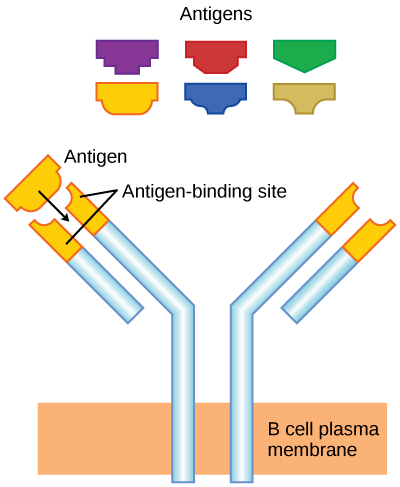
Cada célula B tiene solo un tipo de receptor de antígeno, lo que hace que cada célula B sea diferente. Una vez que las células B maduran en la médula ósea, migran a los ganglios linfáticos u otros órganos linfáticos. Cuando una célula B se encuentra con el antígeno que se une a su receptor, la molécula de antígeno se introduce en la célula por endocitosis y reaparece en la superficie de la célula unida a una molécula MHC de clase II. Cuando se completa este proceso, la célula B se sensibiliza. En la mayoría de los casos, la célula B sensibilizada debe entonces encontrarse con un tipo específico de célula T, llamada célula T auxiliar, antes de que se active. La célula T auxiliar ya debe haber sido activada a través de un encuentro con el antígeno (discutido a continuación).
La célula T auxiliar se une al complejo antígeno-MHC clase II y se induce a liberar citocinas que inducen a la célula B a dividirse rápidamente, lo que produce miles de células idénticas (clonales). Estas células hijas se convierten en células plasmáticas o células B de memoria. Las células B de memoria permanecen inactivas en este punto, hasta que otro encuentro posterior con el antígeno, provocado por una reinfección por la misma bacteria o virus, da como resultado que se dividan en una nueva población de células plasmáticas. Las células plasmáticas, por otro lado, producen y secretan grandes cantidades, hasta 100 millones de moléculas por hora, de moléculas de anticuerpos. Un anticuerpo, también conocido como inmunoglobulina (Ig), es una proteína que es producida por las células plasmáticas después de la estimulación por un antígeno. Los anticuerpos son los agentes de la inmunidad humoral. Los anticuerpos se presentan en la sangre, en las secreciones gástricas y mucosas, y en la leche materna. Los anticuerpos en estos fluidos corporales pueden unirse a patógenos y marcarlos para su destrucción por los fagocitos antes de que puedan infectar las células.
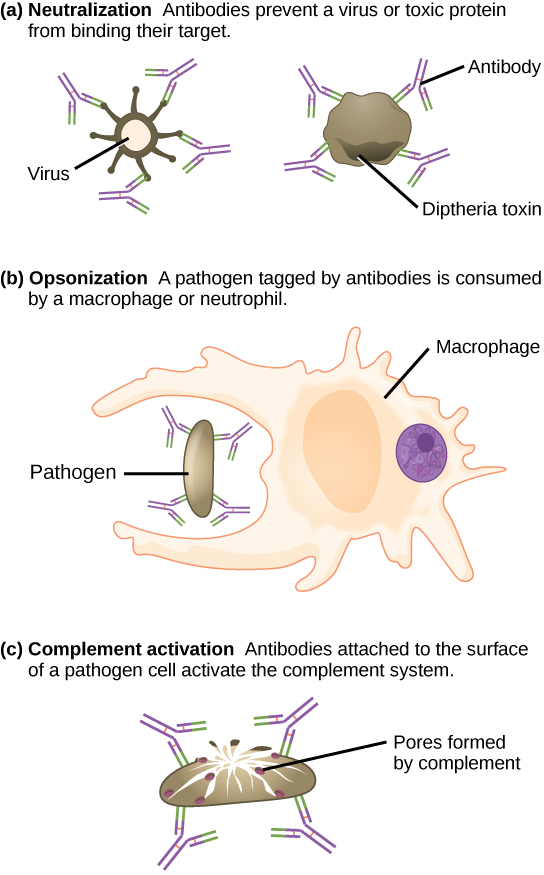
Estos anticuerpos circulan en el torrente sanguíneo y el sistema linfático y se unen con el antígeno cada vez que se encuentra. La unión puede combatir la infección de varias maneras. Los anticuerpos pueden unirse a virus o bacterias e interferir con las interacciones químicas necesarias para que infecten o se unan a otras células. Los anticuerpos pueden crear puentes entre diferentes partículas que contienen sitios antigénicos que los agrupan y evitan su correcto funcionamiento. El complejo antígeno-anticuerpo estimula el sistema del complemento descrito anteriormente, destruyendo la célula portadora del antígeno. Las células fagocíticas, como las ya descritas, son atraídas por los complejos antígeno-anticuerpo, y la fagocitosis se potencia cuando los complejos están presentes. Finalmente, los anticuerpos estimulan la inflamación, y su presencia en el moco y en la piel previene el ataque de patógenos.
Los anticuerpos recubren patógenos extracelulares y los neutralizan bloqueando sitios clave en el patógeno que potencian su infectividad (como los receptores que “acoplan” patógenos en las células hospedadoras) (Figura\(\PageIndex{3}\)). La neutralización de anticuerpos puede evitar que los patógenos entren e infecten las células hospedadoras. Los patógenos neutralizados recubiertos con anticuerpos pueden ser filtrados por el bazo y eliminados en orina o heces.
Los anticuerpos también marcan patógenos para su destrucción por células fagocíticas, como macrófagos o neutrófilos, en un proceso llamado opsonización. En un proceso llamado fijación del complemento, algunos anticuerpos proporcionan un lugar para que las proteínas del complemento se unan. La combinación de anticuerpos y complemento promueve la rápida eliminación de patógenos.
La producción de anticuerpos por las células plasmáticas en respuesta a un antígeno se denomina inmunidad activa y describe la respuesta activa del sistema inmunitario del huésped a una infección o a una vacunación. También hay una respuesta inmune pasiva donde los anticuerpos provienen de una fuente externa, en lugar de las propias células plasmáticas del individuo, y se introducen en el huésped. Por ejemplo, los anticuerpos que circulan en el cuerpo de una mujer embarazada se mueven a través de la placenta hacia el feto en desarrollo. El niño se beneficia de la presencia de estos anticuerpos hasta varios meses después del nacimiento. Además, una respuesta inmune pasiva es posible inyectando anticuerpos en un individuo en forma de antiveneno a una toxina mordida de serpiente o anticuerpos en el suero sanguíneo para ayudar a combatir una infección de hepatitis. Esto da protección inmediata ya que el cuerpo no necesita el tiempo requerido para montar su propia respuesta.
Inmunidad mediada por células
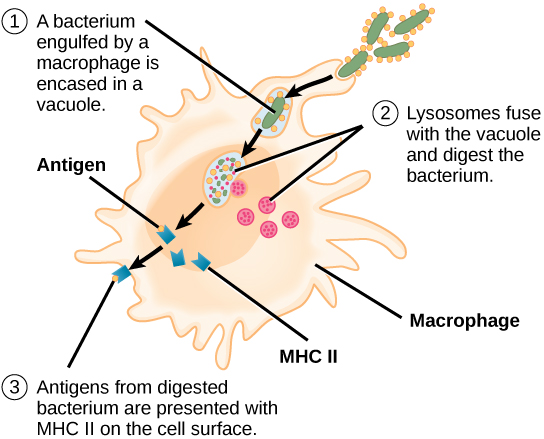
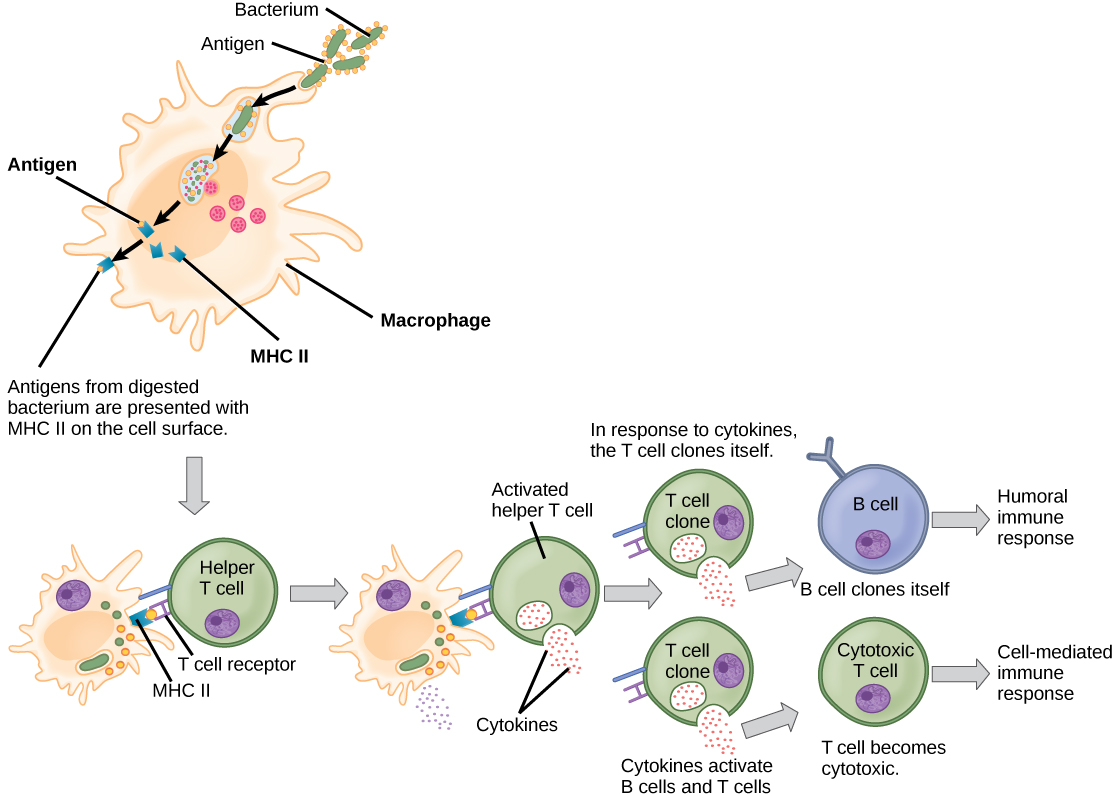
A diferencia de las células B, los linfocitos T son incapaces de reconocer patógenos sin ayuda. En cambio, las células dendríticas y los macrófagos primero engullen y digieren patógenos en cientos o miles de antígenos. Luego, una célula presentadora de antígeno (APC) detecta, envuelve e informa la respuesta inmune adaptativa sobre una infección. Cuando se detecta un patógeno, estos APC lo engullirán y descompondrán a través de fagocitosis. Los fragmentos de antígeno serán transportados entonces a la superficie de la APC, donde servirán como indicador a otras células inmunes. Una célula dendrítica es una célula inmune que limpia materiales antigénicos en su entorno y los presenta en su superficie. Las células dendríticas se localizan en la piel, los revestimientos de la nariz, los pulmones, el estómago y los intestinos. Estas posiciones son lugares ideales para encontrar patógenos invasores. Una vez que son activados por patógenos y maduran para convertirse en APC, migran al bazo o a un ganglio linfático. Los macrófagos también funcionan como APC. Después de la fagocitosis por un macrófago, la vesícula fagocítica se fusiona con un lisosoma intracelular. Dentro del fagolisosoma resultante, los componentes se descomponen en fragmentos; los fragmentos se cargan luego en moléculas MHC clase II y se transportan a la superficie celular para la presentación del antígeno (Figura\(\PageIndex{4}\)). Las células T colaboradoras no pueden responder adecuadamente a un antígeno a menos que se procese e incruste en una molécula MHC de clase II. Las APC expresan MHC clase II en sus superficies, y cuando se combinan con un antígeno extraño, estos complejos señalan a un invasor.
Las células T tienen muchas funciones. Algunos responden a las APC del sistema inmune innato e inducen indirectamente respuestas inmunes liberando citocinas. Otros estimulan a las células B para iniciar la respuesta humoral como se describió anteriormente. Otro tipo de célula T detecta señales de APC y mata directamente a las células infectadas, mientras que algunas están involucradas en la supresión de reacciones inmunitarias inapropiadas a antígenos inofensivos o “propios”.
Existen dos tipos principales de células T: los linfocitos T cooperadores (T H) y los linfocitos T citotóxicos (T C). Los linfocitos T H funcionan indirectamente para informar a otras células inmunes sobre posibles patógenos. Los linfocitos T H reconocen antígenos específicos presentados por los complejos MHC clase II de APC. Hay dos poblaciones de células T H: T H 1 y T H 2. Las células T H 1 secretan citoquinas para potenciar las actividades de los macrófagos y otras células T. Las células T H 2 estimulan a las células B sin tratamiento previo para secretar anticuerpos. El que se desarrolle una respuesta inmune T H 1 o una T H 2 depende de los tipos específicos de citocinas secretadas por las células del sistema inmunitario innato, lo que a su vez depende de la naturaleza del patógeno invasor.
Las células T citotóxicas (T C) son el componente clave de la parte mediada por células del sistema inmune adaptativo y atacan y destruyen las células infectadas. Las células T C son particularmente importantes en la protección contra infecciones virales; esto se debe a que los virus se replican dentro de las células donde están protegidos del contacto extracelular con anticuerpos circulantes. Una vez activada, la T C crea un gran clon de células con un conjunto específico de receptores de superficie celular, como en el caso de la proliferación de células B activadas. Al igual que con las células B, el clon incluye células T C activas y células T C de memoria inactivas. Las células T C activas resultantes identifican entonces las células hospedadoras infectadas. Debido al tiempo requerido para generar una población de células T y B clonales, existe un retraso en la respuesta inmune adaptativa en comparación con la respuesta inmune innata.
Las células T C intentan identificar y destruir las células infectadas antes de que el patógeno pueda replicarse y escapar, deteniendo así la progresión de las infecciones intracelulares. Las células T C también apoyan a los linfocitos NK para destruir los cánceres tempranos. Las citocinas secretadas por la respuesta T H 1 que estimula los macrófagos también estimulan las células T C y potencian su capacidad para identificar y destruir células infectadas y tumores. En la Figura aparece un resumen de cómo se activan las respuestas inmunitarias humorales y mediadas por células\(\PageIndex{5}\).
Las células plasmáticas B y las células T C se denominan colectivamente células efectoras porque están involucradas en “efectuar” (provocar) la respuesta inmune de matar patógenos y células hospedadoras infectadas.
Memoria Inmunológica
El sistema inmune adaptativo tiene un componente de memoria que permite una respuesta rápida y grande a la reinvasión del mismo patógeno. Durante la respuesta inmune adaptativa a un patógeno que no se había encontrado antes, conocida como la respuesta inmune primaria, las células plasmáticas secretoras de anticuerpos y las células T diferenciadas aumentan, luego se estabilizan con el tiempo. A medida que las células B y T maduran en células efectoras, un subconjunto de las poblaciones vívidas se diferencia en células B y T de memoria con las mismas especificidades de antígeno (Figura\(\PageIndex{6}\)). Una célula de memoria es un linfocito B o T específico de antígeno que no se diferencia en una célula efectora durante la respuesta inmune primaria, pero que puede convertirse inmediatamente en una célula efectora tras la reexposición al mismo patógeno. A medida que la infección se elimina y los estímulos patógenos desaparecen, los efectores ya no son necesarios y sufren apoptosis. En contraste, las células de memoria persisten en la circulación.
CONEXIÓN ARTE
El antígeno Rh se encuentra en los glóbulos rojos Rh positivos. Una hembra Rh negativa generalmente puede llevar un feto Rh positivo a término sin dificultad. Sin embargo, si tiene un segundo feto Rh positivo, su cuerpo puede lanzar un ataque inmune que cause enfermedad hemolítica del recién nacido. ¿Por qué cree que la enfermedad hemolítica es solo un problema durante el segundo embarazo o posteriores?
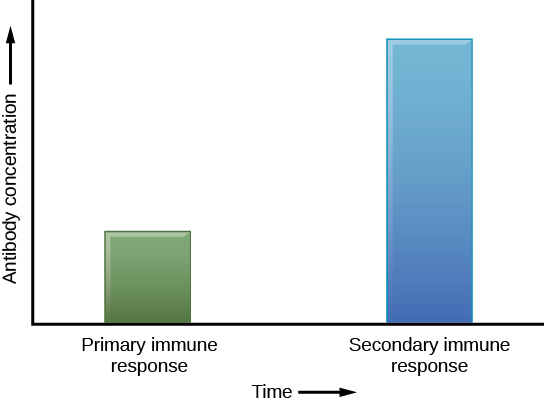
Si el patógeno nunca se vuelve a encontrar durante la vida del individuo, las células de memoria B y T circularán por algunos años o incluso varias décadas y morirán gradualmente, nunca habiendo funcionado como células efectoras. Sin embargo, si el hospedador se vuelve a exponer al mismo tipo de patógeno, las células de memoria circulantes se diferenciarán inmediatamente en células plasmáticas y células T C sin entrada de APC o células T H. Esto se conoce como la respuesta inmune secundaria. Una razón por la que la respuesta inmune adaptativa se retrasa es porque lleva tiempo identificar, activar y proliferar las células B y T sin tratamiento previo con las especificidades antigénicas apropiadas. En la reinfección, se omite este paso, y el resultado es una producción más rápida de defensas inmunitarias. Las células B de memoria que se diferencian en células plasmáticas producen cantidades de anticuerpos de decenas a cien veces mayores que las que se secretaron durante la respuesta primaria (Figura\(\PageIndex{7}\)). This rapid and dramatic antibody response may stop the infection before it can even become established, and the individual may not realize they had been exposed.
La vacunación se basa en el conocimiento de que la exposición a antígenos no infecciosos, derivados de patógenos conocidos, genera una respuesta inmune primaria leve. La respuesta inmune a la vacunación puede no ser percibida por el huésped como enfermedad, pero aún así confiere memoria inmune. Cuando se expone al patógeno correspondiente al que se vacunó un individuo, la reacción es similar a una exposición secundaria. Debido a que cada reinfección genera más células de memoria y mayor resistencia al patógeno, algunos cursos de vacunas involucran una o más vacunas de refuerzo para imitar exposiciones repetidas.
El Sistema Linfático
La linfa es el líquido acuoso que baña tejidos y órganos y contiene glóbulos blancos protectores pero no contiene eritrocitos. La linfa se mueve alrededor del cuerpo a través del sistema linfático, que se compone de vasos, conductos linfáticos, glándulas linfáticas y órganos, como amígdalas, adenoides, timo y bazo.
Aunque el sistema inmunitario se caracteriza por la circulación de células por todo el cuerpo, la regulación, maduración e intercomunicación de factores inmunes ocurren en sitios específicos. La sangre hace circular células inmunitarias, proteínas y otros factores a través del cuerpo. Aproximadamente 0.1 por ciento de todas las células en la sangre son leucocitos, que incluyen monocitos (el precursor de macrófagos) y linfocitos. La mayoría de las células en la sangre son glóbulos rojos. Las células del sistema inmune pueden viajar entre los distintos sistemas linfático y circulatorio sanguíneo, los cuales están separados por el espacio intersticial, por un proceso llamado extravasación (que pasa a través del tejido circundante).
Recordemos que las células del sistema inmune se originan a partir de células madre en la médula ósea. La maduración de las células B ocurre en la médula ósea, mientras que las células progenitoras migran de la médula ósea y se desarrollan y maduran en células T vívidas en el órgano llamado timo.
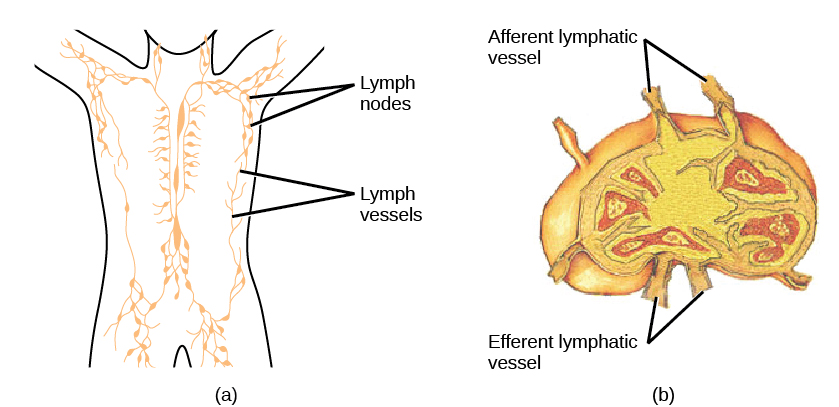
En la maduración, los linfocitos T y B circulan a diversos destinos. Los ganglios linfáticos dispersos por todo el cuerpo albergan grandes poblaciones de células T y B, células dendríticas y macrófagos (Figura\(\PageIndex{8}\)). La linfa reúne antígenos a medida que drena de los tejidos. Estos antígenos luego se filtran a través de los ganglios linfáticos antes de que la linfa vuelva a la circulación. Las APC en los ganglios linfáticos capturan y procesan antígenos e informan a los linfocitos cercanos sobre posibles patógenos.
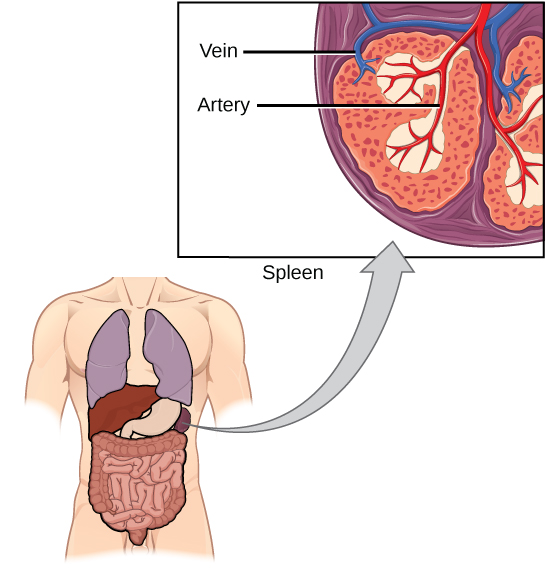
El bazo alberga células B y T, macrófagos, células dendríticas y células NK (Figura\(\PageIndex{9}\)). El bazo es el sitio donde las APC que tienen partículas extrañas atrapadas en la sangre pueden comunicarse con los linfocitos. Los anticuerpos son sintetizados y secretados por células plasmáticas activadas en el bazo, y el bazo filtra sustancias extrañas y patógenos complejados con anticuerpos de la sangre. Funcionalmente, el bazo es a la sangre como lo son los ganglios linfáticos a la linfa.
Sistema Inmune Mucosa
Las respuestas inmunes innatas y adaptativas componen el sistema inmune sistémico (afectando a todo el cuerpo), que es distinto del sistema inmune mucoso. El tejido linfoide asociado a la mucosa (MALT) es un componente crucial de un sistema inmune funcional porque las superficies mucosas, como las fosas nasales, son los primeros tejidos sobre los que se depositan patógenos inhalados o ingeridos. El tejido mucoso incluye la boca, la faringe y el esófago, y los tractos gastrointestinal, respiratorio y urogenital.
La inmunidad mucosa está formada por MALT, que funciona independientemente del sistema inmune sistémico, y que tiene sus propios componentes innatos y adaptativos. MALT es una colección de tejido linfático que se combina con el tejido epitelial que recubre la mucosa en todo el cuerpo. Este tejido funciona como barrera inmune y respuesta en áreas del cuerpo con contacto directo con el ambiente externo. Los sistemas inmunitarios sistémicos y mucosos utilizan muchos de los mismos tipos de células. Las partículas extrañas que llegan a MALT son absorbidas por células epiteliales absorbentes y entregadas a APC localizadas directamente debajo del tejido mucoso. Las APC del sistema inmune mucoso son principalmente células dendríticas, con células B y macrófagos que tienen papeles menores. Los antígenos procesados que se muestran en las APC son detectados por las células T en el MALT y en las amígdalas, adenoides, apéndice o los ganglios linfáticos mesentéricos del intestino. Las células T activadas luego migran a través del sistema linfático y al sistema circulatorio a los sitios mucosos de infección.
Tolerancia Inmune
El sistema inmunológico tiene que ser regulado para evitar respuestas derrochadoras e innecesarias a sustancias inofensivas, y lo que es más importante, para que no se atente al “yo”. La capacidad adquirida para prevenir una respuesta inmune innecesaria o dañina a una sustancia extraña detectada que se sabe que no causa enfermedad, o autoantígenos, se describe como tolerancia inmune. El mecanismo principal para desarrollar la tolerancia inmune a los autoantígenos ocurre durante la selección de células débilmente autoenlazantes durante la maduración de los linfocitos T y B. Existen poblaciones de células T que suprimen la respuesta inmune a los autoantígenos y que suprimen la respuesta inmune después de que la infección se haya aclarado para minimizar el daño de las células hospedadoras inducido por la inflamación y lisis celular. La tolerancia inmune está especialmente bien desarrollada en la mucosa del sistema digestivo superior debido a la tremenda cantidad de sustancias extrañas (como las proteínas de los alimentos) que encuentran las APC de la cavidad oral, la faringe y la mucosa gastrointestinal. La tolerancia inmune es provocada por APC especializadas en el hígado, los ganglios linfáticos, el intestino delgado y el pulmón que presentan antígenos inofensivos a una población diversa de células T reguladoras (T reg), linfocitos especializados que suprimen la inflamación local e inhiben la secreción de inmunidad estimuladora factores. El resultado combinado de las células T reg es prevenir la activación inmunológica y la inflamación en compartimentos tisulares no deseados y permitir que el sistema inmunitario se centre en los patógenos.
Resumen de la Sección
La respuesta inmune adaptativa es una respuesta de acción más lenta, más duradera y más específica que la respuesta innata. Sin embargo, la respuesta adaptativa requiere información del sistema inmune innato para funcionar. Las APC muestran antígenos en moléculas MHC a células T sin tratamiento previo. Las células T con receptores de superficie celular que se unen a un antígeno específico se unirán a esa APC. En respuesta, las células T se diferencian y proliferan, convirtiéndose en células T H o células T. Las células T H estimulan las células B que han engullido y presentado antígenos derivados de patógenos. Las células B se diferencian en células plasmáticas que secretan anticuerpos, mientras que las células T C destruyen las células infectadas o cancerosas. Las células de memoria son producidas por células B y T activadas y proliferantes y persisten después de una exposición primaria a un patógeno. Si se produce una reexposición, las células de memoria se diferencian en células efectoras sin la entrada del sistema inmune innato. El sistema inmune mucoso es en gran medida independiente del sistema inmune sistémico pero funciona en paralelo para proteger las extensas superficies mucosas del cuerpo. La tolerancia inmune es provocada por las células T reg para limitar las reacciones a antígenos inofensivos y a las moléculas propias del cuerpo.
Conexiones de arte
Figura\(\PageIndex{6}\): El antígeno Rh se encuentra en glóbulos rojos Rh positivos. Una hembra Rh negativa generalmente puede llevar un feto Rh positivo a término sin dificultad. Sin embargo, si tiene un segundo feto Rh positivo, su cuerpo puede lanzar un ataque inmune que cause enfermedad hemolítica del recién nacido. ¿Por qué cree que la enfermedad hemolítica es solo un problema durante el segundo embarazo o posteriores?
- Contestar
-
Si la sangre de la madre y el feto se mezcla, las células de memoria que reconocen el antígeno Rh del feto pueden formarse en la madre al final del primer embarazo. Durante embarazos posteriores, estas células de memoria lanzan un ataque inmune sobre las células sanguíneas fetales de un feto Rh positivo. La inyección de anticuerpo anti-RH durante el primer embarazo evita que se produzca la respuesta inmune.
Glosario
- inmunidad activa
- una inmunidad que se produce como resultado de la actividad de las propias células del cuerpo y no de anticuerpos adquiridos de una fuente externa
- inmunidad adaptativa
- una respuesta inmune específica que ocurre después de la exposición a un antígeno ya sea de un patógeno o de una vacunación
- anticuerpo
- una proteína que es producida por las células plasmáticas después de la estimulación por un antígeno; también conocida como inmunoglobulina
- antígeno
- una macromolécula que reacciona con las células del sistema inmune y que puede o no tener un efecto estimulante
- célula presentadora de antígeno (APC)
- una célula inmune que detecta, envuelve e informa la respuesta inmune adaptativa sobre una infección presentando el antígeno procesado en su superficie celular
- Célula B
- un linfocito que madura en la médula ósea
- respuesta inmune mediada por células
- una respuesta inmune adaptativa que es controlada por células T
- linfocitos T citotóxicos (T C)
- una célula inmune adaptativa que mata directamente las células infectadas a través de enzimas y que libera citocinas para mejorar la respuesta inmune
- células dendríticas
- una célula inmune que procesa material antigénico y lo presenta en la superficie de su célula en moléculas MHC clase II e induce una respuesta inmune en otras células
- célula efectora
- un linfocito que se ha diferenciado, como una célula B, una célula plasmática o una célula T citotóxica
- linfocitos T auxiliares (T H)
- una célula del sistema inmune adaptativo que se une a las APC a través de moléculas MHC clase II y estimula las células B o secreta citocinas para iniciar la respuesta inmune
- respuesta inmune humoral
- la respuesta inmune adaptativa que es controlada por células B activadas y anticuerpos
- tolerancia inmune
- una capacidad adquirida para prevenir una respuesta inmune innecesaria o dañina a un cuerpo extraño detectado que se sabe que no causa enfermedad
- linfa
- el líquido acuoso presente en el sistema circulatorio linfático que baña tejidos y órganos con glóbulos blancos protectores y no contiene eritrocitos
- celda de memoria
- un linfocito B o T específico de antígeno que no se diferencia en una célula efectora durante la respuesta inmune primaria pero que puede convertirse inmediatamente en una célula efectora tras la reexposición al mismo patógeno
- molécula de clase mayor de histocompatibilidad (MHC) II
- una proteína que se encuentra en la superficie de las células presentadoras de antígeno que indica a las células inmunitarias si la célula es normal o está infectada o cancerosa; proporciona el molde apropiado en el que los antígenos pueden cargarse para su reconocimiento por los linfocitos
- inmunidad pasiva
- una inmunidad que no resulta de la actividad de las propias células inmunitarias del cuerpo sino de la transferencia de anticuerpos de un individuo a otro
- respuesta inmune primaria
- la respuesta del sistema inmune adaptativo a la primera exposición a un antígeno
- respuesta inmune secundaria
- la respuesta del sistema inmune adaptativo a una segunda o posterior exposición a un antígeno mediado por células de memoria
- Célula T
- un linfocito que madura en la glándula del timo