
2.4: Estructura y Función- Proteínas II
- Page ID
- 52887

Fuente: BiochemFFA_2_3.pdf. Todo el libro de texto está disponible gratuitamente de los autores en http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy
En esta sección, esperamos dar vida a la conexión entre estructura y función de las proteínas. Hasta el momento, hemos descrito características notables de los cuatro elementos (primario, secundario, terciario y cuaternario) de la estructura proteica y discutimos ejemplos de proteínas/motivos que los exhiben. En esta sección, examinaremos desde una perspectiva funcional algunas proteínas/dominios cuya función se basa en la estructura secundaria, terciaria o cuaternaria. Por supuesto, es un enfoque un poco estrecho atribuir la función proteica a cualquier componente de la estructura, pero nuestra esperanza es al presentar estos ejemplos, podemos dar vida a la forma en que la estructura secundaria, terciaria y cuaternaria de una proteína conduce a las funciones que tiene.
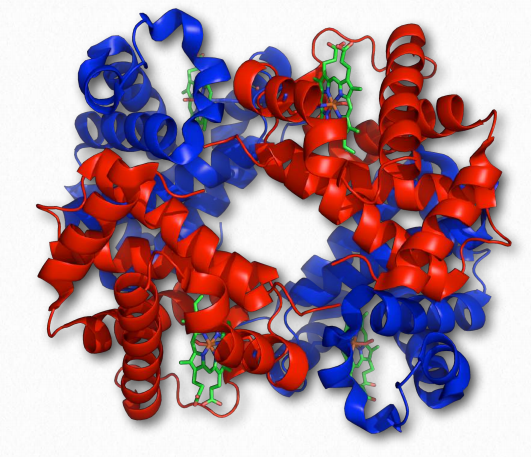
Hemoglobina Wikipedia
Proteínas fibrosas - estructura secundaria
Las proteínas cuyos papeles celulares o extracelulares tienen un fuerte componente estructural están compuestas principalmente de estructura primaria y segunda, con poco plegamiento de las cadenas. Así, tienen muy poca estructura terciaria y son de naturaleza fibrosa. Las proteínas que exhiben estos rasgos son comúnmente insolubles en agua y se denominan proteínas fibrosas (también llamadas escleroproteínas). Los ejemplos descritos en esta categoría se encuentran exclusivamente en animales donde desempeñan papeles en carne, tejidos conectivos y estructuras externas endurecidas, como el pelo. También contienen las tres estructuras proteicas fibrosas comunes α-hélices (queratinas), β-hilos/láminas (fibroína y elastina) y triples hélices (colágeno). Las proteínas fibrosas tienen cierta similitud de secuencia de aminoácidos. Cada uno posee una abundancia de secuencias repetitivas de aminoácidos con pequeños grupos laterales no reactivos. Muchos contienen repeticiones cortas de secuencias, a menudo con glicina.
Queratinas

Las queratinas son una familia de proteínas animales relacionadas que toman numerosas formas. Las α-queratinas son componentes estructurales de la capa externa de la piel humana y son integrales al cabello, uñas, garras, plumas, picos, escamas y pezuñas. Las queratinas proporcionan fuerza a los tejidos, como la lengua, y más de 50 queratinas diferentes están codificadas en el genoma humano. A nivel celular, las queratinas comprenden los filamentos intermedios del citoesqueleto. Las queratinas α contienen principalmente hélices α-pero también pueden tener estructuras β-fila/lámina. Las hélices α-individuales a menudo se entrelazan para formar bobinas de estructuras enrolladas y estas hebras también se pueden unir adicionalmente mediante enlaces disulfuro, lo que aumenta considerablemente la resistencia estructural. Esto es particularmente relevante para la α-queratina en el cabello, que contiene cerca de 14% de cisteína. El olor del cabello quemado y el de los químicos utilizados para rizar/desenrizar el cabello (romper/rehacer enlaces disulfuro) surgen de sus componentes sulfurosos. Las β-queratinas están compuestas por β-láminas, como su nombre lo indica.
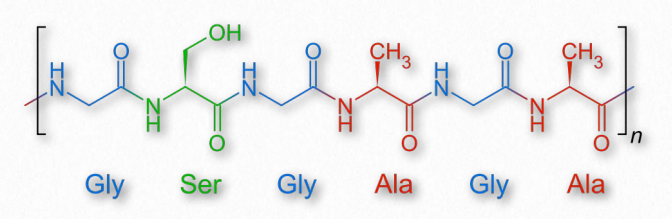
Fibroína
Proteína fibrosa insoluble que es un componente de la seda de las arañas y las larvas de polillas y otros insectos, la fibroína se compone de hebras β antiparalelas apretadas juntas para formar láminas β-. La estructura primaria de la fibroína es una secuencia repetitiva corta con glicina en cada resto (Figura 2.57). Los pequeños grupos R de la glicina y alanina en la secuencia repetitiva permiten el empaquetamiento apretado característico de las fibras de seda. Enlace de Wikipedia AQUÍ Elastina Como sugiere su nombre, la elastina es una proteína con características elásticas que funciona en muchos tejidos del cuerpo para permitirles retomar sus formas después de expandirse o contraerse. La proteína es rica en glicina y prolina y puede comprender más del 50% del peso de las arterias secas y desgrasadas.

Elastina
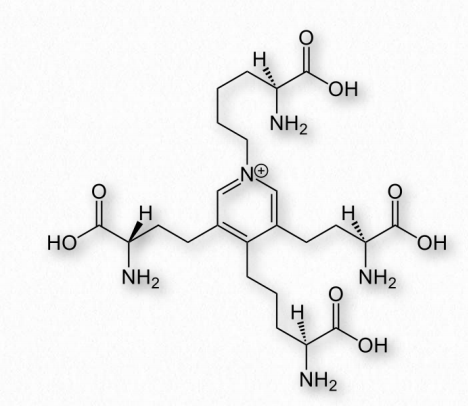
se elabora uniendo proteínas de tropoelastina a través de residuos de lisina para hacer un complejo duradero reticulado por desmosina. En las arterias, la elastina ayuda con la propagación de la onda de presión para facilitar el flujo sanguíneo.
Colágeno
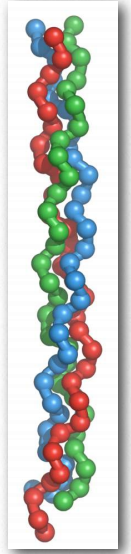
El colágeno es la proteína más abundante en los mamíferos, ocupando hasta un tercio de la masa total. Hay al menos 16 tipos de colágeno. Sus fibras son un componente importante de los tendones y también se encuentran abundantemente en la piel. El colágeno también es prominente en la córnea, cartílago, hueso, vasos sanguíneos y el intestino.
La estructura del colágeno es un ejemplo de una hélice de hélices, estando compuesta por tres cadenas helicoidales izquierdas que cada una se enrollan juntas de manera diestra para hacer la fibra de colágeno (Figura 2.60). Cada hélice se estira más que una hélice α, dándole una apariencia extendida. En el interior de la estructura de triple hélice, solo se encuentran restos de glicina, ya que las cadenas laterales de otros aminoácidos son demasiado voluminosas. Las cadenas de colágeno tienen la estructura repetitiva glycine-n donde m es a menudo prolina y n es a menudo hidroxiprolina (Figura 2.61).

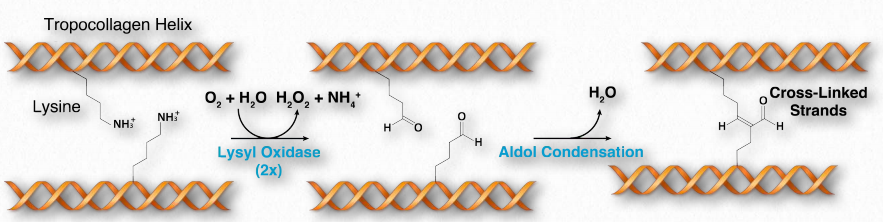
El colágeno se sintetiza en forma de pre-procolágeno. El procesamiento del preprocolágeno en el retículo endoplásmico da como resultado glicosilación, eliminación de la secuencia 'pre' e hidroxilación de residuos de lisina y prolina (ver abajo). Los hidróxidos pueden formar reticulaciones covalentes entre sí, fortaleciendo las fibras de colágeno. A medida que el procolágeno se exporta fuera de la célula, las proteasas lo recortan, dando como resultado una forma final de colágeno llamada tropocolágeno.
Hidroxilación
La hidroxilación de las cadenas laterales de prolina y lisina ocurre postraduccionalmente en una reacción catalizada por prolil-4-hidroxilasa y lisil-hidroxilasa (lisil oxidasa), respectivamente. La reacción requiere vitamina C. Dado que la hidroxilación de estos residuos es esencial para la formación de triples hélices estables a temperatura corporal, la deficiencia de vitamina C da como resultado colágeno débil e inestable y, en consecuencia, tejidos conectivos debilitados. Es la causa de la enfermedad conocida como escorbuto. El colágeno hidrolizado se utiliza para elaborar gelatina, lo cual es importante en la industria alimentaria. colágenos. Enlace de Wikipedia AQUÍ
Laminas
Las laminas son proteínas fibrosas que proporcionan estructura en el núcleo celular y desempeñan un papel en la regulación de la transcripción. Son similares a las proteínas que componen los filamentos intermedios, pero tienen aminoácidos adicionales en una bobina de la proteína. Las laminas ayudan a formar la lámina nuclear en el interior de la envoltura nuclear y desempeñan un papel importante en el ensamblaje y desmontaje de esta última en el proceso de mitosis. También ayudan a posicionar los poros nucleares. En el proceso de mitosis, el desmontaje de la envoltura nuclear es promovido por la fosforilación de las laminas por una proteína llamada factor promotor de la mitosis y el ensamblaje se ve favorecido al revertir la reacción (desfosforilación).
Dominios estructurales - estructura terciaria
Toda proteína globular se basa en su estructura terciaria para realizar su función, así que en lugar de tratar de encontrar proteínas representativas para la estructura terciaria (¡una tarea casi imposible!) , nos enfocamos aquí en algunos elementos de estructura terciaria que son comunes a muchas proteínas. Estos son los dominios estructurales y difieren de los motivos estructurales de estructura supersecundaria por ser más grandes (25-500 aminoácidos), tener una secuencia de aminoácidos conservada, y un historial de evolución y funcionamiento independientemente de las cadenas proteicas en las que se encuentran. Los dominios estructurales son unidades fundamentales de estructura terciaria y se encuentran en más de una proteína. Un dominio estructural es autoestabilizante y a menudo se pliega independientemente del resto de la cadena proteica.
Cremallera de leucina
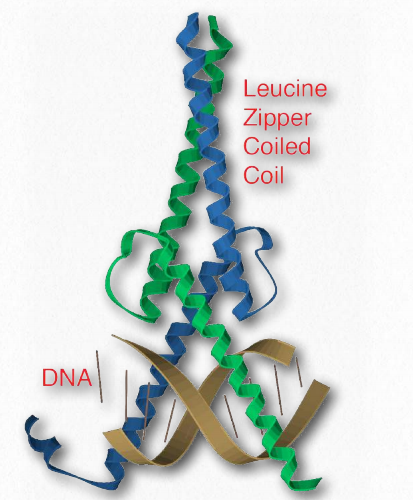
Una característica común de muchas proteínas eucariotas de unión a ADN, las cremalleras de leucina se caracterizan por un conjunto repetitivo de residuos de leucina en una proteína que interactúan como una cremallera para favorecer la dimerización. Otra parte del dominio tiene aminoácidos (comúnmente arginina y lisina) que le permiten interactuar con la doble hélice del ADN (Figura 2.63). Los factores de transcripción que contienen cremalleras de leucina incluyen Jun-B, CREB y AP-1 fos/ jun.
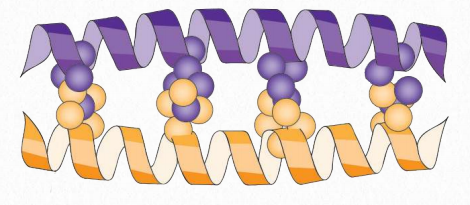
Dedos de zinc
Los dominios estructurales más cortos son los dedos de zinc, que obtienen su nombre por el hecho de que uno o más iones de zinc coordinados estabilizan su estructura similar a un dedo. A pesar de su nombre, algunos dedos de zinc no se unen al zinc. Existen muchos dominios estructurales clasificados como dedos de zinc y estos se agrupan en diferentes familias. Los dedos de zinc se identificaron por primera vez como componentes de los factores de transcripción de unión al ADN, pero ahora se sabe que otros se unen a estructuras de ARN, proteínas e incluso lípidos. Las cadenas laterales de cisteína e histidina suelen desempeñar un papel en la coordinación del zinc.
Dominio Src SH 2
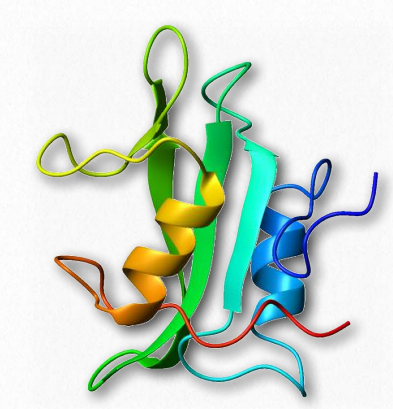
La oncoproteína Src contiene un dominio estructural SH 2 conservado que reconoce y se une a las cadenas laterales de tirosina fosforiladas en otras proteínas (Figura 2.65). La fosforilación es una actividad fundamental en la señalización y fosforilación de tirosina y la interacción entre proteínas portadoras de señales es críticamente necesaria para la comunicación celular. El dominio SH 2 se encuentra en más de 100 proteínas humanas.
Dominio de hélice girada-hélice

La hélice-vuelta-hélice es un dominio común que se encuentra en las proteínas de unión al ADN, que consiste en dos hélices α-separadas por un pequeño número de aminoácidos. Como se ve en la Figura 2.66, las partes de hélice del dominio estructural interactúan con las bases en el surco mayor del ADN. Las hélices α-individuales en una proteína son parte de una estructura de hélice-giro-hélice, donde el giro separa las hélices individuales.
Dominio de homología de Pleckstrin
Los dominios de homología de Pleckstrin (PH) son dominios proteicos con funciones importantes en el proceso de señalización. Esto surge en parte de la afinidad por la unión de inositidos fosforilados, como PIP2 y PIP3, que se encuentra en la Figura 2.66 - Dominio Hélice-Turn-Hélice de una Proteína Unida al ADN Wikipedia Figura 2.65 - Dominio SH 2 Wikipedia membranas biológicas. Los dominios PH también pueden unirse a proteínas G y proteína quinasa C. El dominio abarca alrededor de aminoácidos y se encuentra en numerosas proteínas de señalización. Estas incluyen Akt/Rac Serina/Treonina Proteína Quinasas, BTK/LTK/Tec tirosina proteína quinasas, sustrato receptor de insulina (IRS-1), fosfolipasa C específica de fosfatidilinositoly varias proteínas de levadura involucradas en la regulación del ciclo celular.
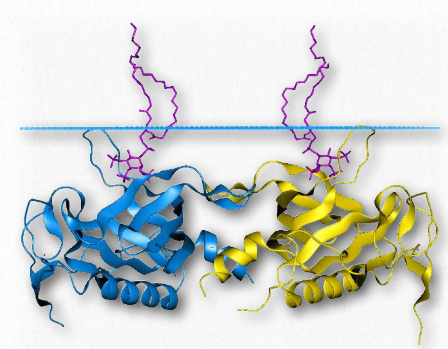
Proteínas globulares estructurales
Las enzimas catalizan reacciones y proteínas como la hemoglobina realizan importantes funciones especializadas. La selección evolutiva ha reducido y eliminado los desechos para que podamos estar seguros de que cada proteína en una célula tiene una función, aunque en algunos casos tal vez no sepamos qué es. En ocasiones la estructura de la ProFigura 2.68 - Relación de la membrana basal con el epitelio, endotelio, y teína del tejido conectivo es su función primaria porque la estructura proporciona estabilidad, organización, conexiones otras propiedades importantes. Es con esto en mente que presentamos las siguientes proteínas.
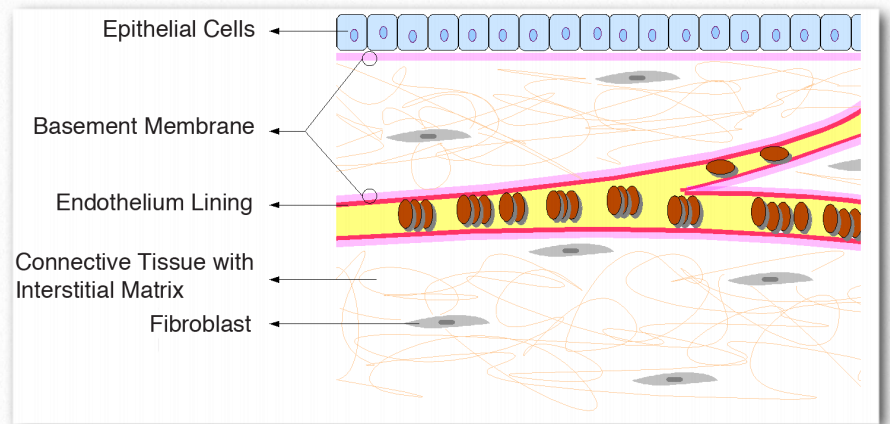
Membrana del sótano
La membrana basal es una matriz extracelular estratificada de tejido compuesta por fibras proteicas (colágeno tipo IV) y glicosaminoglicanos que separa el epitelio de otros tejidos (Figura 2.68). Más importante aún, la membrana basal actúa como un pegamento para mantener unidos los tejidos. La piel, por ejemplo, está anclada al resto del cuerpo por la membrana basal. Las membranas basales proporcionan una interfaz de interacción entre las células y el entorno que las rodea, facilitando así los procesos de señalización. Desempeñan papeles en la diferenciación durante la embriogénesis y también en el mantenimiento de la función en organismos adultos.
Actina
La actina es la proteína globular más abundante que se encuentra en la mayoría de los tipos de células eucariotas, que comprende hasta el 20% del peso de las células musculares. Se han identificado proteínas similares en bacterias (MReb) y arqueones (Ta0583). La actina es una subunidad monomérica capaz de polimerizar fácilmente en dos tipos diferentes de filamentos. Los microfilamentos son componentes principales del citoesqueleto y son actuados por la miosina en la contracción de las células musculares (Ver AQUÍ). Actina se discutirá con más detalle en la siguiente sección AQUÍ.
Filamentos Intermedios
Los filamentos intermedios forman parte del citoesqueleto en muchas células animales y están compuestos por más de 70 proteínas diferentes. Se les llama intermedios porque su tamaño (diámetro promedio = 10 nm) se encuentra entre el de los microfilamentos (7 nm) y los microtúbulos (25 nm).
Los componentes intermedios del filamento incluyen proteínas fibrosas, como las queratinas y las laminas, que son nucleares, así como formas citoplásmicas. Los filamentos intermedios dan flexibilidad a las células debido a sus propias propiedades físicas. Pueden, por ejemplo, estirarse a varias veces de su longitud original.
Seis tipos
Hay seis tipos diferentes de filamentos intermedios. Los tipos I y II son ácidos o básicos y se atraen entre sí para hacer filamentos más grandes. Incluyen queratinas epiteliales y queratinas tricocíticas (componentes capilares). Las proteínas de tipo III incluyen cuatro proteínas estructurales: desmina, GFAP (proteína ácida fibrilar glial), periferina y vimentina. El tipo IV también es un agrupamiento de tres proteínas y una estructura multiproteica (neurofilamentos). Las tres proteínas son α-internexina, sinemina y sincoilina. Los filamentos intermedios tipo V abarcan las laminas, las cuales dan estructura al núcleo. La fosforilación de las laminas conduce a su desmontaje y esto es importante en el proceso de mitosis. La categoría Tipo VI incluye solo una única proteína conocida como nestin.
Tubulina
Un tercer tipo de filamento que se encuentra en las células es el de las microbutulas. Comprendidos por un polímero de dos unidades de una proteína globular llamada tubulina, los microtúbulos proporcionan “rieles” para que las proteínas motoras muevan orgánulos y otra “carga” de una parte de una célula a otra. Los microtúbulos y la tubulina se discuten con más detalle AQUÍ.
Vimentín
La vimentina (Figura 2.70) es la proteína más ampliamente distribuida de los filamentos intermedios. Se expresa en fibroblastos, leucocitos y células endoteliales de los vasos sanguíneos. La proteína tiene un papel importante manteniendo la posición de los orgánulos en el citoplasma, con uniones al núcleo, mitocondrias y retículo endoplásmico (Figura 2.70). La vimentina aporta elasticidad a las células y resiliencia que no surge de los microtúbulos o microfilamentos. Los ratones heridos que carecen del gen de la vimentina sobreviven, pero tardan más en sanar las heridas que los ratones de tipo salvaje. Vimentina también controla el movimiento del colesterol desde los lisosomas hasta el sitio de esterificación. El resultado es una reducción en la cantidad de colesterol almacenado dentro de las células y tiene implicaciones para las células suprarrenales, las cuales deben tener ésteres de colesterol.
Mucín
Las mucinas son un grupo de proteínas que se encuentran en el tejido epitelial animal que tienen muchos residuos de glicosil en ellas y típicamente son de alto peso molecular (1 a 10 millones de Da). Son de tipo gel en su carácter y a menudo se utilizan para la lubricación. El moco está compuesto por mucinas. Además de la lubricación, las mucinas también ayudan a controlar la mineralización, como la formación ósea en organismos vertebrados y la calcificación en equinodermos. También juegan un papel en el sistema inmunológico al ayudar a unir patógenos. Las mucinas se secretan comúnmente en las superficies mucosas (fosas nasales, ojos, boca, oídos, estómago, genitales, ano) o en fluidos, como la saliva. Debido a su extensa mucosilación, las mucinas contienen una cantidad considerable de agua (dándoles la sensación “viscosa”) y son resistentes a la proteólisis.
Vinculina
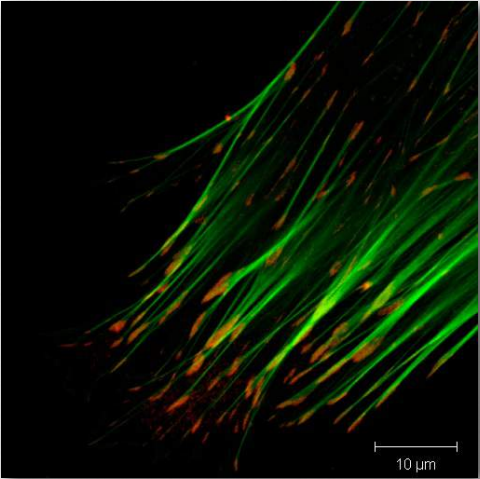
La vinculina (Figura 2.72) es una proteína citoesquelética de membrana que se encuentra en las estructuras de adhesión focal de células de mamífero. Se encuentra en las uniones célula-célula y célula-matriz e interactúa con integrinas, talina, paxilinas y F-actina. Se cree que la vinculina ayuda (junto con otras proteínas) en el anclaje de microfilamentos de actina a la membrana (Figura 2.71). La unión de vinculina a actina y a talina se regula mediante polifosfoinosítidos y puede ser inhibida por fosfolípidos ácidos.
Sindecanos
Los sindecanos son proteínas transmembrana que hacen un solo paso con una larga cadena de aminoácidos (24-25 residuos) a través de las membranas plasmáticas y facilitan la interacción de los receptores acoplados a proteínas G con la Figura 2.71 - Filamentos de actina (verdes) unidos a la vinculina en ligandos de Wikipedia de adhesión focal (rojo), como el crecimiento factores, fibronectina, colágenos (I, III y IV) y antitrombina-1. Los sindecanos suelen tener 3-5 cadenas de sulfato de heparano y sulfato de condroitina unidas a ellos.
El sulfato de heparán se puede escindir en el sitio de una herida y estimular la acción del factor de crecimiento de fibroblastos en el proceso de curación. El papel de los sindecanos en la adhesión célula-célula se muestra en células mutantes que carecen de sindecano I que no se adhieren bien entre sí. También se sabe que Syndecan 4 se adhiere a la integrina. Los sindecanos también pueden inhibir la propagación de tumores por la capacidad del ectodominio sindecano 1 para suprimir el crecimiento de las células tumorales sin afectar a las células epiteliales normales.
Defensina
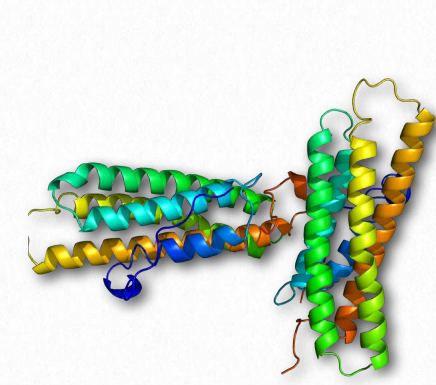
Las defensinas (Figura 2.73) son un grupo de pequeñas proteínas catiónicas (ricas en residuos de cisteína) que sirven como péptidos de defensa del huésped en organismos vertebrados e invertebrados. Protegen contra la infección por diversas bacterias, hongos y virus. Las defensinas contienen entre 18 y 45 aminoácidos con (típicamente) aproximadamente 6-8 residuos de cisteína. En el sistema inmune, las defensinas ayudan a matar las bacterias envueltas por la fagocitosis por las células epiteliales y los neutrófilos. Matan 120 Figura 2.72 - Vinculina Wikipedia bacterias actuando como ionóforos - uniéndose a la membrana y abriendo estructuras en forma de poros para liberar iones y nutrientes de las células.
Adhesiones focales
En la célula, las adherencias focales son estructuras que contienen múltiples proteínas que enlazan mecánicamente las estructuras citoesqueléticas (haces de actina) con la matriz extracelular. Son dinámicas, con proteínas trayendo y saliendo con señales respecto al ciclo celular, motilidad celular, y más casi constantemente. Las adherencias focales sirven como anclajes y como centro de señalización en ubicaciones celulares donde las integrinas se unen a moléculas y donde ocurren eventos de agrupamiento de membranas. Más de 100 proteínas diferentes se encuentran en las adherencias focales.
Las adherencias focales comunican mensajes importantes a las células, actuando como sensores para actualizar la información sobre el estado de la matriz extracelular, lo que, a su vez, ajusta/afecta sus acciones. En las células sedentarias, son más estables que en las células en movimiento porque cuando las células se mueven, los contactos focales de adhesión se establecen en el “frente” y se retiran en la parte trasera a medida que avanza el movimiento. Esto puede ser muy importante en la capacidad de los glóbulos blancos para encontrar daño tisular.
Ankyrin
Las anquirinas (Figura 2.74) son una familia de proteínas adaptadoras de membrana que sirven como “anclajes” para interconectar proteínas integrales de membrana con el citoesqueleto de membrana de espectrina-actina. Las anquirinas se anclan a la membrana plasmática mediante palmitoil-CoA covalentemente enlazada. Se unen a la subunidad β de la espectrina y al menos a una docena de grupos de proteínas integrales de membrana. Las proteínas anquirinas contienen cuatro dominios funcionales: una región N-terminal con 24 repeticiones de anquirina en tándem, un dominio central de unión a espectrinas, un “dominio de muerte” que interactúa con proteínas apoptóticas y un dominio regulador C-terminal que varía significativamente entre diferentes anquirinas.
Spectrin

La espectrina (Figuras 2.75 y 2.76) es una proteína del citoesqueleto celular que juega un papel importante en el mantenimiento de su estructura y la integridad de la membrana plasmática. En los animales, la espectrina da forma a los glóbulos rojos. La espectrina se encuentra dentro de la capa interna de la membrana plasmática eucariota donde forma una red de arreglos pentagonales o hexagonales.
Las fibras de espectrina se acumulan juntas en complejos de unión de actina y también se unen a la anquirina para mayor estabilidad, así como a numerosas proteínas integrales de membrana, como la glicoforina.
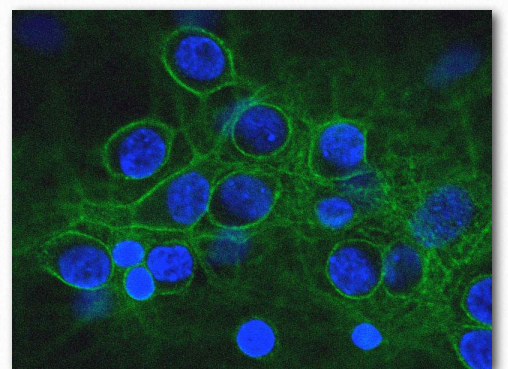
Integrinas
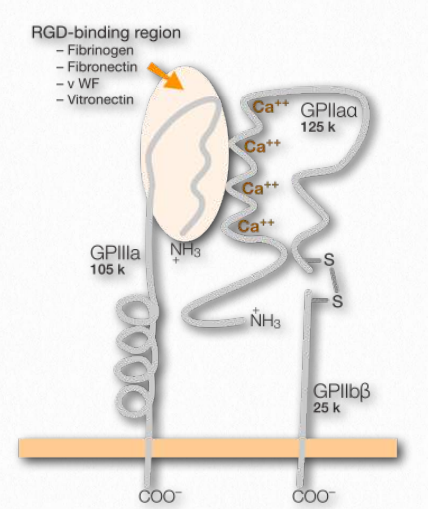
En los organismos multicelulares, las células necesitan conexiones, tanto entre sí como con la matriz extracelular. Facilitando estas uniones en el extremo celular están las proteínas transmembrana conocidas como integrinas (Figura 2.77). Las integrinas se encuentran en todas las células metazoicas. Los ligandos para las integrinas incluyen colágeno, fibronectina, laminina y vitronectina. Las integrinas funcionan no solo en la unión, sino también en la comunicación, la migración celular, los enlaces virales (adenovirus, por ejemplo) y la coagulación de la sangre. Las integrinas son capaces de detectar señales químicas y mecánicas sobre la matriz extracelular y mover esa información a dominios intracelulares como parte del proceso de transducción de señales. Dentro de las células, las respuestas a las señales afectan la forma celular, la regulación del ciclo celular, el movimiento o los cambios en otros receptores celulares en la membrana. El proceso es dinámico y permite respuestas rápidas según sea necesario, por ejemplo en el proceso de coagulación de la sangre, donde la integrina conocida como GPIbIIIa (en la superficie de las plaquetas sanguíneas) se une a la fibrina en un coágulo a medida que se desarrolla.
Las integrinas funcionan junto con otros receptores, incluyendo inmunoglobulinas, otras moléculas de adhesión celular, cadherinas, selectinas y sindecanos. En los mamíferos las proteínas tienen un gran número de subunidades -18 cadenas α- y 8 β-cadenas. Son un puente entre sus enlaces fuera de la célula con la matriz extracelular (ECM) y sus enlaces dentro de la célula con el citoesqueleto. Las integrinas desempeñan un papel central en la formación y estabilidad de las adherencias focales. Se trata de grandes complejos moleculares que surgen de la agrupación de conexiones integrina-ECM. En el proceso de movimiento celular, las integrinas en el “frente” de la célula (en la dirección del movimiento), hacen nuevas fijaciones al sustrato y liberan conexiones al sustrato en la parte posterior de la célula. Estas últimas integrinas son entonces endocitosadas y reutilizadas.
Las integrinas también ayudan a modular la transducción de señales a través de receptores de tirosina quinasa en la membrana celular regulando el movimiento de los adaptadores a la membrana plasmática. La integrina β1c, por ejemplo, recluta la fosfatasa Shp2 al receptor del factor de crecimiento de insulina para hacer que se desfosforile, volviéndose así fuera de la señal que comunica. Las integrinas también pueden ayudar a reclutar moléculas de señalización dentro de la célula a tirosina quinasas activadas para ayudarles a comunicar sus señales.
Cadherinas

Las cadherinas (Figura 2.78) constituyen una clase tipo 1 de proteínas transmembrana que desempeñan un papel importante en la adhesión celular. Requieren iones calcio para funcionar, formando uniones adherentes que mantienen unidos los tejidos (Ver Figura 2.69). Las células de un tipo específico de cadherina se agruparán preferentemente entre sí en preferencia a asociarse con células que contienen un tipo de cadherina diferente. Las caderinas son tanto receptores como lugares para que los ligandos se unan. Ayudan en el posicionamiento adecuado de las células en el desarrollo, separación de diferentes capas de tejido y migración celular.
Selectins
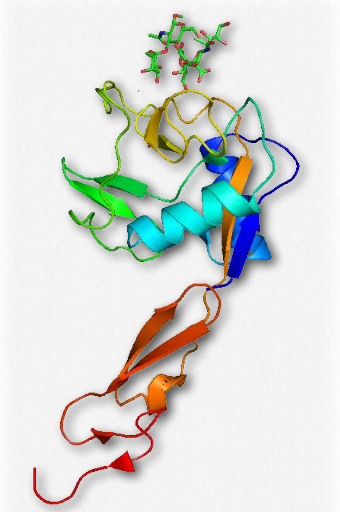
Las selectinas (Figura 2.79) son glicoproteínas de adhesión celular que se unen a moléculas de azúcar. Como tal, son un tipo de lectina -proteínas que se unen a polímeros de azúcar (ver AQUÍ también). Todas las selectinas tienen un dominio de lectina dependiente de calcio N-terminal, un solo dominio transmembrana y una cola citoplásmica intracelular.
Existen tres tipos diferentes de selectinas, 1) E-selectina (endotelial); 2) L (linfocítica; y 3) P (plaquetas y células endoteliales. Las selectinas funcionan en el homing de linfocitos (adhesión de linfocitos sanguíneos a células de órganos linfoides), en procesos inflamatorios y en metástasis de cáncer. Cerca del sitio de inflamación, la P-selectina en la superficie de las células capilares sanguíneas interactúa con glicoproteínas en las superficies celulares leucocitarias. Esto tiene el efecto de ralentizar el movimiento del leucocito. En el sitio diana de la inflamación, la E-selectina en las células endoteliales del vaso sanguíneo y la L-selectina en la superficie del leucocito se unen a sus respectivos carbohidratos, deteniendo el movimiento leucocitario. El leucocito cruza entonces la pared del capilar y comienza la respuesta inmune. Las selectinas están involucradas en los procesos inflamatorios de asma, psoriasis, escleroris múltiple y artritis reumatoide.
Lamininas
Las lamininas son glicoproteínas de la matriz extracelular que son componentes principales de la lámina basal y afectan la diferenciación celular, la migración y la adhesión. Se secretan a la matriz extracelular donde se incorporan y son esenciales para el mantenimiento y supervivencia de los tejidos. Cuando las lamininas son defectuosas, los músculos pueden no formarse adecuadamente y dar lugar a distrofia muscular.
Las lamininas están asociadas con fibronectina, entactina y proteínas perlecanas en redes de colágeno tipo IV y se unen a receptores de integrina en la membrana plasmática. Como consecuencia, las lamininas contribuyen a la unión celular, diferenciación, forma y movimiento. Las proteínas son triméricas en estructura, teniendo una cadena α, una cadena β y una cadena γ. Se conocen quince combinaciones de diferentes cadenas.
Vitronectina
La vitronectina es una glicoproteína (75kDa) que se encuentra en el suero sanguíneo (plaquetas), la matriz extracelular y en el hueso. Promueve el proceso de adhesión y propagación celular y se une a varios inhibidores de proteasa (serpinas). Se secreta de las células y se cree que juega un papel en la coagulación de la sangre y la malignidad de los tumores. Un dominio de la vitronectina se une al inhibidor activador del plasminógeno y actúa para estabilizarlo. Otro dominio de la proteína se une a las proteínas de integrina celular, como el receptor de vitronectina que ancla las células a la matriz extracelular.
Cateninas
Las cateninas son una familia de proteínas que interactúan con las proteínas cadherinas en la adhesión celular (Figura 2.69). Se conocen cuatro tipos principales de cateninas, α-, β-, γ- y δ-catenina. Las cateninas juegan un papel en la organización celular antes de que ocurra el desarrollo y ayudan a regular el crecimiento celular. α-catenina y β-catenina se encuentran en uniones adherentes con cadherina y ayudan a las células a mantener las capas epiteliales. Las cadherinas están conectadas a los filamentos de actina del citoesqueleto y las cateninas juegan el papel crítico. Las cateninas son importantes para el proceso mediante el cual la división celular se inhibe cuando las células entran en contacto entre sí (inhibición por contacto).
Cuando los genes de catenina están mutados, las adherencias celulares de cadherina pueden desaparecer y puede resultar tumorigénesis. Se ha encontrado que las cateninas están asociadas con cáncer colorrectal y muchas otras formas de cáncer.
Glicóforinas
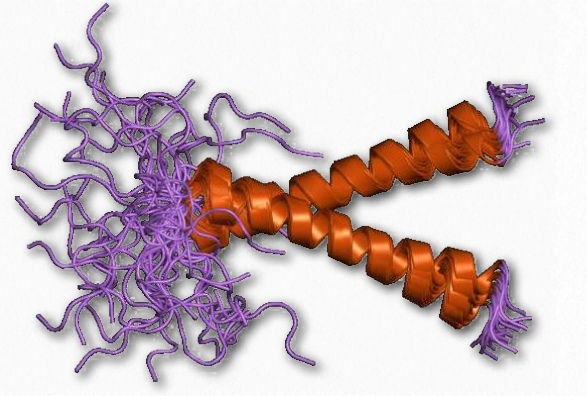
Todas las proteínas de membrana descritas hasta ahora son notables por las conexiones que hacen con otras proteínas y estructuras celulares. Algunas proteínas de membrana, sin embargo, están diseñadas para reducir las conexiones celulares con proteínas de otras células. Esto es particularmente importante para las células sanguíneas donde la “adherencia” es indeseable excepto en lo que respecta a la coagulación.
Las glicoforinas (Figura 2.80) son sialoglicoproteínas que abarcan la membrana de los glóbulos rojos. Están fuertemente glicosiladas (60%) .y ricas en ácido siálico, dando a las células una capa muy hidrofílica (y cargada negativamente), lo que les permite circular en el torrente sanguíneo sin adherirse a otras células o a las paredes de los vasos.
Se han identificado cinco glicoforinas, cuatro (A, B, C y D) de membranas aisladas y una quinta forma (E) de codificación en el genoma humano. Las proteínas son abundantes, formando alrededor del 2% del total de proteínas de membrana en estas células. Las glicoforinas tienen un papel importante en la regulación de las propiedades mecánicas y la forma de la membrana de los glóbulos Debido a que algunas glicoforinas pueden expresarse en diversos tejidos no eritroides (particularmente Glicoforina C), la importancia de sus interacciones con el esqueleto de la membrana puede tener una significación biológica considerable.
Cooperatividad y alosterismo - estructura cuaternaria
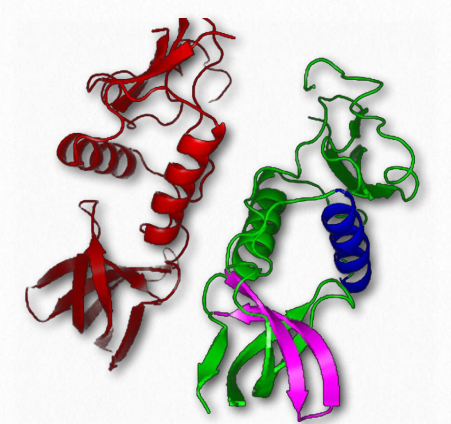
La estructura cuaternaria, por supuesto, describe las interacciones de subunidades individuales de una proteína multisubunidad (Figura 2.81). El resultado de estas interacciones puede dar lugar a importantes fenómenos biológicos, como la unión cooperativa de sustratos a una proteína y efectos alostéricos sobre la acción de una enzima.
Los efectos alostéricos pueden ocurrir por una serie de mecanismos, pero una característica común es que la unión de un efector a una subunidad enzimática causa (o bloquea) la enzima en un Tstate (menos actividad) o un estado R (más actividad). Los efectores pueden ser sustratos enzimáticos (efectores homotrópicos) o no sustratos (efectores heterotrópicos). El alosterismo se cubrirá con más profundidad en el capítulo Catálisis AQUÍ.
Comenzamos nuestra consideración de la estructura cuaternaria con una discusión sobre la cooperatividad, cómo surge en la hemoglobina de la proteína multisubunidad y cómo sus propiedades contrastan con las de la proteína mioglobina de subunidad única relacionada.
Cooperatividad
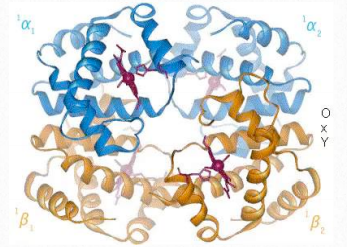
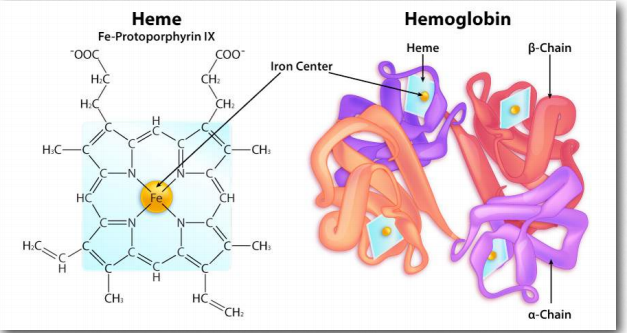
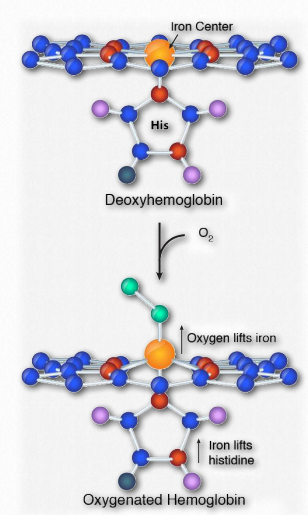
La cooperatividad se define como el fenómeno donde la unión de una molécula ligando por una proteína favorece la unión de moléculas adicionales del mismo tipo. La hemoglobina, por ejemplo, exhibe cooperatividad cuando la unión de una molécula de oxígeno por el hierro del grupo hemo en una de las cuatro subunidades provoca un ligero cambio de conformación en la subunidad. Esto sucede porque el hierro hemo está unido a una cadena lateral de histidina y la unión del oxígeno 'levanta' el hierro junto con el anillo de histidina (también conocido como el anillo de imidazol).
Película 2.3 - Cambios estructurales de la hemoglobina en la unión de oxígeno Wikipedia
Dado que cada subunidad de hemoglobina interactúa con las otras subunidades e influye en ellas, también son inducidas a cambiar de forma ligeramente cuando la primera subunidad se une al oxígeno (una transición que se describe como que va del estado T al estado R). Estos cambios de forma favorecen también a cada una de las subunidades restantes que se unen al oxígeno. Esto es muy importante en los pulmones donde el oxígeno es captado por la hemoglobina, ya que la unión de la primera molécula de oxígeno facilita la rápida absorción de más moléculas de oxígeno. En los tejidos, donde la concentración de oxígeno es menor, el oxígeno sale de la hemoglobina y las proteínas voltean del estado R de nuevo al Tstate.
Transporte de CO 2
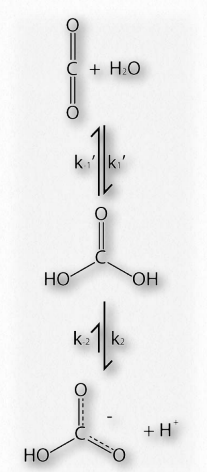
La cooperatividad es solo uno de los muchos aspectos estructurales fascinantes de la hemoglobina que ayudan al cuerpo a recibir oxígeno donde se necesita y recogerlo donde sea abundante. La hemoglobina también ayuda en el transporte del producto de la respiración celular (dióxido de carbono) desde los tejidos que la producen hasta los pulmones donde se exhala. Al igual que la unión del oxígeno a la hemoglobina, la unión de otras moléculas a la hemoglobina afecta su afinidad por el oxígeno. El efecto es particularmente pronunciado al comparar las características de unión al oxígeno de las cuatro subunidades de hemoglobina con la unión al oxígeno de la subunidad única de la proteína mioglobina relacionada (Figura 2.83).
Diferente unión de oxígeno
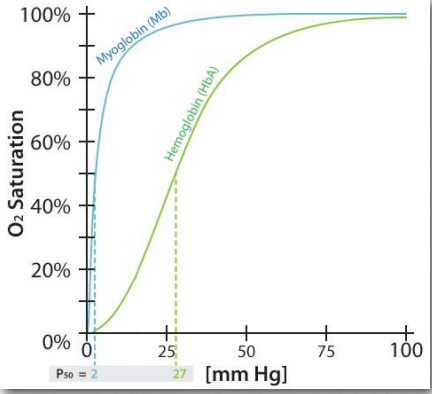
Al igual que la hemoglobina, la mioglobina contiene un hierro en un grupo hemo que se une al oxígeno. La estructura de la proteína globina en la mioglobina es muy similar a la estructura de las globinas en la hemoglobina y se cree que la hemoglobina evolucionó a partir de la mioglobina en la historia evolutiva. Como se observa en la Figura 2.83, la curva de unión de la hemoglobina para el oxígeno es en forma de S (sigmoidal), mientras que la curva de unión para la mioglobina es hiperbólica. Lo que esto nos dice es que la afinidad de la hemoglobina por el oxígeno es baja a una baja concentración de oxígeno, pero aumenta a medida que aumenta la concentración de oxígeno. Dado que la mioglobina se satura muy rápidamente de oxígeno, incluso bajo bajas concentraciones de oxígeno, dice que su afinidad por el oxígeno es alta y no cambia.
Debido a que la mioglobina tiene una sola subunidad, la unión de oxígeno por esa subunidad no puede afectar a ninguna otra subunidad, ya que no hay otras subunidades a las que afectar. En consecuencia, la cooperatividad requiere de más de una subunidad. Por lo tanto, la hemoglobina puede exhibir cooperatividad, pero la mioglobina no, vale la pena señalar que el simple hecho de tener múltiples subunidades no significa que exista cooperatividad. La hemoglobina es una proteína que exhibe la característica, pero muchas proteínas multisubunitarias no.
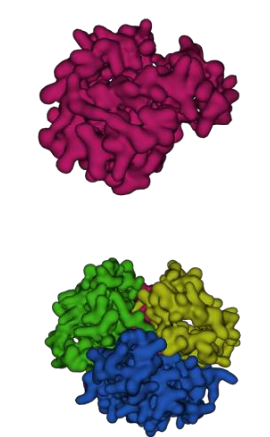
Interactivo 2.2 - Hemoglobina en presencia (arriba) y ausencia (abajo) de oxígeno
Almacenamiento vs. entrega
La falta de capacidad de la mioglobina para ajustar su afinidad por el oxígeno de acuerdo con la concentración de oxígeno (baja afinidad a baja concentración de oxígeno, como en los tejidos y alta afinidad a alta concentración de oxígeno, como en los pulmones) significa que es más adecuada para almacenar oxígeno que para administrarlo de acuerdo a las diferentes necesidades de oxígeno y cuerpo animal. Como veremos, además de la cooperatividad, la hemoglobina tiene otras características estructurales que le permiten entregar oxígeno precisamente donde más se necesita en el cuerpo.
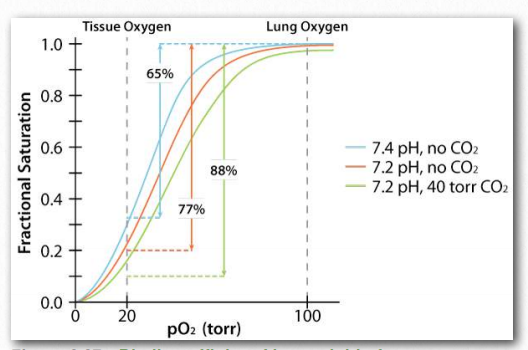
Efecto Bohr
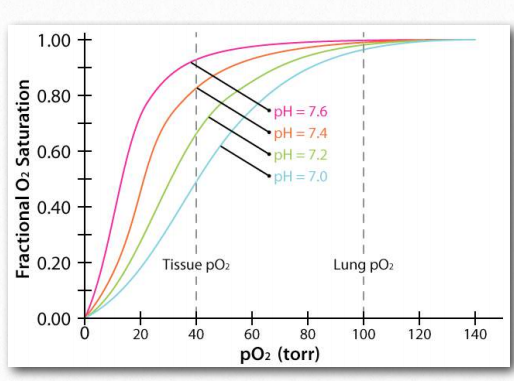
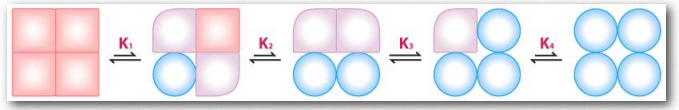
El Efecto Bohr fue descrito por primera vez hace más de 100 años por Christian Bohr, padre del famoso físico, Niels Bohr. Se muestra gráficamente (Figuras 2.86, 2.87 y 2.88), el efecto observado es que la afinidad de la hemoglobina por el oxígeno disminuye a medida que disminuye el pH y a medida que aumenta la concentración de dióxido de carbono. Unión de los protones y dióxido de carbono por amino Figura 2.85 - Modelo secuencial de unión. El modelo secuencial es una forma de explicar la cooperatividad de la hemoglobina. Los cuadrados no representan oxígeno unido. Los círculos representan subunidades unidas con oxígeno y las subunidades redondeadas corresponden a unidades cuya afinidad por el oxígeno aumenta al interactuar con una subunidad que tiene oxígeno unido. Imagen de Aleia Kim Las cadenas laterales ácidas en las proteínas globinas ayudan a facilitar los cambios estructurales en ellas. Más comúnmente, el aminoácido afectado por los protones es la histidina #146 de las cadenas β. Cuando esto sucede, la histidina ionizada puede formar un enlace iónico con la cadena lateral del ácido aspártico #94, lo que tiene el efecto de estabilizar el estado T (estado de unión de oxígeno reducido) y liberar oxígeno. Otras histidinas y la amina de los aminoácidos amino terminales en las cadenas α también son sitios de unión para protones.
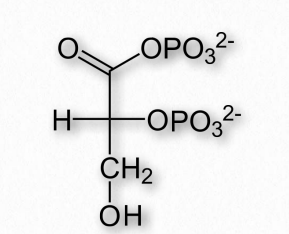
2,3-BPG
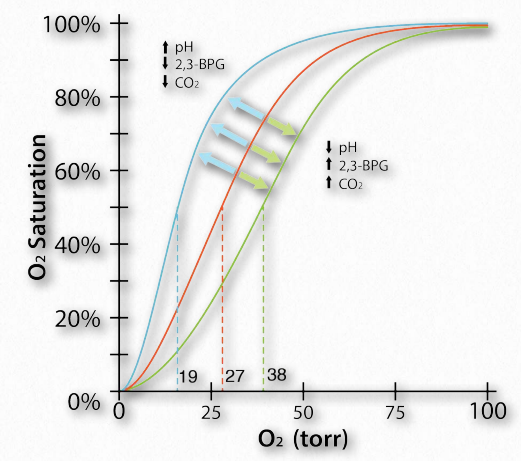
Otra molécula que favorece la liberación de oxígeno por la hemoglobina es el 2,3- bisfosfosglicerato (también llamado 2,3-BPG o simplemente BPG - Figura 2.89). Al igual que los protones y el dióxido de carbono, el 2,3-BPG se produce respirando activamente los tejidos, como subproducto del metabolismo de la glucosa. El mol de 2,3-BPG encaja en el 'agujero de la rosquilla' de la hemoglobina adulta (Figura 2.89). Dicha unión de 2,3-BPG favorece el estado T (unión apretada - baja en oxígeno) de la hemoglobina, que tiene una afinidad reducida por el oxígeno. En ausencia de 2,3-BPG, la hemoglobina puede existir más fácilmente en el estado R (relajado - mayor unión al oxígeno), que tiene una alta afinidad por el oxígeno.
Fumadores
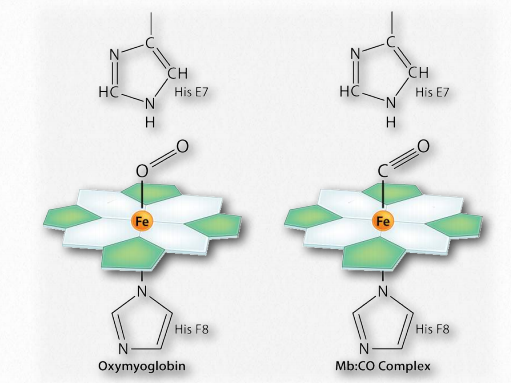
Notablemente, la sangre de los fumadores es mayor en la concentración de 2,3-BPG que los no fumadores, por lo que más de su hemoglobina permanece en el estado T y por lo tanto la capacidad de transporte de oxígeno de los fumadores es menor que la de los no fumadores.Otra razón por la que la capacidad de carga de oxígeno de los fumadores es menor que la de los no fumadores es que el humo del cigarrillo contiene monóxido de carbono y esta molécula, que tiene dimensiones casi idénticas al oxígeno molecular, efectivamente supera con el oxígeno por unirse al átomo de hierro del hemo (Figura 2.90). Parte de la toxicidad del monóxido de carbono se debe a su capacidad para unirse a la hemoglobina y evitar que el oxígeno se una.
Dióxido de carbono
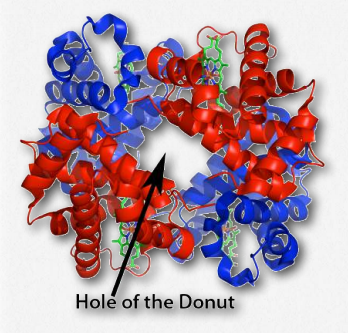
El dióxido de carbono se une para formar un carbamato cuando se une a la α-amina de cada cadena de globina. El proceso de formación de esta estructura libera un protón, lo que ayuda a mejorar aún más el efecto Bohr. Fisiológicamente, la unión de CO2 y H+ tiene importancia porque los tejidos que respiran activamente (como los músculos que se contraen) requieren oxígeno y liberan protones y dióxido de carbono. Cuanto mayor sea la concentración de protones y dióxido de carbono, más oxígeno se libera para alimentar a los tejidos que más lo necesitan.
Alrededor del 40% de los protones liberados y aproximadamente el 20% del dióxido de carbono son transportados de regreso a los pulmones por la hemoglobina. El resto viaja como parte del sistema de amortiguación de bicarbonato o como CO2 disuelto. En los pulmones, el proceso se invierte. Los pulmones tienen un pH más alto que los tejidos respiratorios, por lo que los protones se liberan de la hemoglobina y el CO2 también se libera para ser exhalado.
Hemoglobina fetal
La hemoglobina adulta libera oxígeno cuando se une a 2,3- BPG. Esto contrasta con la hemoglobina fetal, que tiene una configuración ligeramente diferente (α2γ2) que la hemoglobina adulta (α2β2). La hemoglobina fetal tiene una mayor afinidad por el oxígeno que la hemoglobina materna, lo que permite que el feto obtenga oxígeno de manera efectiva de la sangre de la madre. Parte de la razón de la mayor afinidad de la hemoglobina fetal por el oxígeno es que no se une a 2,3-BPG. En consecuencia, la hemoglobina fetal permanece en el estado R mucho más que la hemoglobina adulta y debido a esto, la hemoglobina fetal tiene mayor afinidad por el oxígeno que la hemoglobina adulta y puede alejar el oxígeno de la hemoglobina adulta. Así, el feto puede obtener oxígeno de la madre.
Enfermedad de células falciformes
Las mutaciones en los genes de globina que codifican la hemoglobina a veces pueden tener consecuencias perjudiciales. La anemia falciforme (también llamada anemia falciforme) es una enfermedad de transmisión genética que surge de tales mutaciones. Existen diferentes formas de la enfermedad. Es un rasgo recesivo, es decir, que para ser afligido con él, un individuo debe heredar dos copias del gen mutado.
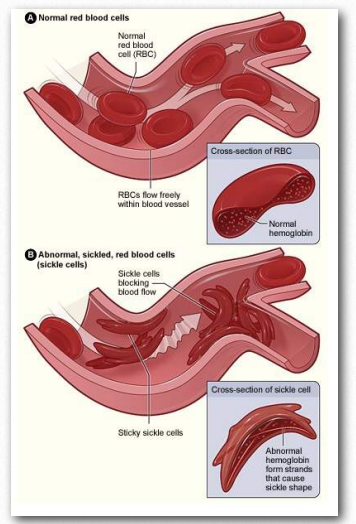
La forma predominante de hemoglobina en adultos es la hemoglobina A, designada HbA (dos cadenas α y dos cadenas β). La forma mutante se conoce como HBs. La mutación más común es una mutación A a T en el centro del codón para el séptimo aminoácido (algunos esquemas de conteo lo llaman el sexto aminoácido) de la cadena β. Esto da como resultado la conversión de un codón GAG a GTG y por lo tanto cambia el aminoácido especificado en esa posición de un ácido glutámico a una valina. Este cambio menor coloca un pequeño parche hidrófobo de aminoácidos en la superficie de las cadenas de β-globina.
La polimerización
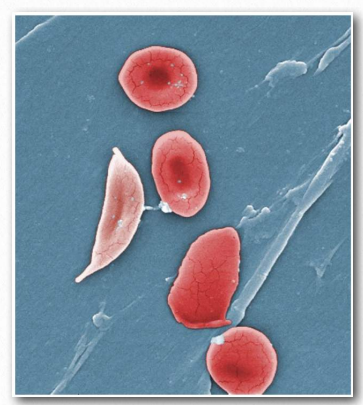
En condiciones de bajo oxígeno, estos parches hidrófobos se asociarán entre sí para hacer polímeros largos de moléculas de hemoglobina. El resultado es que los glóbulos rojos que los contienen cambiarán de forma de ser redondeados a formar la forma de una hoz (Figura 2.94). Los glóbulos rojos redondeados pasan fácilmente a través de pequeños capilares, pero las células falciformes no lo hacen.
Peor aún, bloquean el flujo de otras células sanguíneas. Los tejidos donde ocurren estos bloqueos ya son bajos en oxígeno, por lo que detener el flujo de sangre a través de ellos hace que pasen rápidamente anaeróbicos, causando dolor y, en algunos casos, la muerte del tejido. En circunstancias graves, puede resultar la muerte de glóbulos rojos falciformes. A la enfermedad se le conoce como una anemia debido a que la hoz de los glóbulos rojos se dirige a ellos para su extracción por el sistema de monitoreo de sangre del cuerpo, por lo que una persona con la enfermedad tiene un número reducido crónicamente de glóbulos rojos.
Ventaja heterocigoto
Curiosamente, parece haber una ventaja selectiva para las personas que son heterocigóticas para la enfermedad en áreas donde la malaria es prominente. Los heterocigotos no sufren evidentes efectos nocivos de la enfermedad, pero sus glóbulos rojos parecen ser más susceptibles a la ruptura cuando se infectan. Como consecuencia, el parásito tiene menos posibilidades de reproducirse y la persona infectada tiene mayores posibilidades de supervivencia.
El efecto protector del gen mutante, sin embargo, no se extiende a las personas que padecen la enfermedad en toda regla (homocigotos para el gen mutante). Los tratamientos para la enfermedad incluyen transfusión, manejo del dolor y evitar esfuerzos intensos. El fármaco hidroxiurea se ha relacionado con la reducción en el número y severidad de los ataques, así como un incremento en el tiempo de sobrevivencia1,2. Parece funcionar reactivando la expresión del gen de la hemoglobina fetal, que normalmente no se sintetiza de manera significativa después de aproximadamente 6 semanas de edad.
Atadura de oxígeno
Los animales tienen necesidades de oxígeno que difieren de todos los demás organismos. El oxígeno, por supuesto, es el aceptor de electrones terminal en los animales y es necesario para que el transporte de electrones funcione. Cuando el transporte de electrones está funcionando, la generación de ATP por las células es muchas veces más eficiente que cuando está ausente. Dado que el ATP abundante es esencial para la contracción muscular y los animales se mueven mucho, para atrapar presas, para hacer ejercicio, para escapar del peligro, etc., es importante tener un suministro abundante de oxígeno.
Esto es particularmente una preocupación profunda dentro de los tejidos donde la difusión de oxígeno solo (como ocurre en los insectos) no entrega cantidades suficientes necesarias para la supervivencia a largo plazo. El tema no es un problema para las plantas ya que, en su mayor parte, sus movimientos están en gran parte relacionados con el crecimiento y por lo tanto no tienen necesidades/demandas rápidamente cambiantes de oxígeno que tienen los animales. Los organismos unicelulares tienen una variedad de mecanismos para obtener oxígeno y sobrevivir sin él. Otras dos proteínas importantes que se unen al oxígeno además de la hemoglobina son la mioglobina y la hemocianina.
Mioglobina
La mioglobina es la principal proteína de almacenamiento de oxígeno que se encuentra en los tejidos musculares animales. A diferencia de la hemoglobina, que circula por todo el cuerpo, la proteína mioglobina solo se encuentra en el tejido muscular y aparece en la sangre solo después de una lesión. Al igual que la hemoglobina, la mioglobina se une al oxígeno en un grupo hemo protésico que contiene.
El color rojo de la carne surge del hemo de la mioglobina y el dorado de la carne al cocinarla proviene de la oxidación del ion ferroso (Fe++) del hemo de la mioglobina al ion férrico (Fe+++) vía oxidación en el proceso de cocción. A medida que la carne se asienta en nuestra atmósfera (un ambiente rico en oxígeno), se produce la oxidación de Fe++ a Fe+++, dejando el color marrón señalado anteriormente. Si la carne se almacena en un ambiente de monóxido de carbono (CO), el CO se une al grupo hemo y reduce la cantidad de oxidación, manteniendo la carne con un aspecto rojo por un período de tiempo más largo.
Alta afinidad
La mioglobina (Figura 2.97) muestra mayor afinidad por el oxígeno a bajas concentraciones de oxígeno que la hemoglobina y, por lo tanto, es capaz de absorber el oxígeno suministrado por la hemoglobina bajo estas condiciones. La alta afinidad de la mioglobina por el oxígeno la hace más adecuada para el almacenamiento de oxígeno que para la entrega. La proteína existe como una sola subunidad de globina (a diferencia de la hemoglobina, que contiene cuatro subunidades) y está relacionada con las subunidades que se encuentran en la hemoglobina. Los mamíferos que se sumergen profundamente en el océano, como las ballenas y las focas, tienen músculos con una abundancia particularmente alta de mioglobina. Cuando la concentración de oxígeno en los músculos cae a niveles bajos, la mioglobina libera su oxígeno, funcionando así como una “batería” de oxígeno que entrega combustible de oxígeno cuando es necesario y se aferra a él en todas las demás condiciones. La mioglobina tiene la distinción de ser la primera proteína por la que la estructura 3D fue determinada por cristalografía de rayos X de John Kendrew en 1958, logro por el que más tarde ganó el Premio Nobel.
Hemocianina
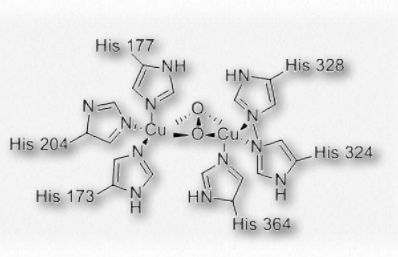
La hemocianina es la proteína que transporta oxígeno en los cuerpos de moluscos y artrópodos. Es una proteína que contiene cobre que no se encuentra dentro de las células sanguíneas de estos organismos, sino que se encuentra suspendida en la hemolinfa circulante que poseen. El sitio de unión al oxígeno de la hemocianina contiene un par de cationes de cobre (I) coordinados directamente a la proteína por los anillos de imidazol de seis cadenas laterales de histidina.

La mayoría, pero no todas las hemocianinas se unen al oxígeno de manera no cooperativa y son menos eficientes que la hemoglobina en el transporte de oxígeno. Notablemente, las hemocianinas de los cangrejos herradura y algunos otros artrópodos, de hecho, unen el oxígeno de manera cooperativa. La hemocianina contiene muchas proteínas subunitarias, cada una con dos átomos de cobre que pueden unirse a una molécula de oxígeno (O2). Las proteínas subunitarias tienen masas atómicas de aproximadamente 75 kilodaltons (kDa). Estos pueden estar dispuestos en dímeros o hexámeros dependiendo de la especie. Las superestructuras compuestas por complejos dímeros o hexámeros están dispuestas en cadenas o agrupaciones y tienen pesos moleculares de más de 1500 kDa.