
2.8: Estructura y Función - Lípidos y Membranas
- Page ID
- 52946

Fuente: BiochemFFA_2_7.pdf. Todo el libro de texto está disponible gratuitamente de los autores en http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy

Los lípidos son un grupo diverso de moléculas que comparten la característica de que al menos una porción de ellas es hidrófoba. Los lípidos juegan muchos papeles en las células, incluyendo servir como almacenamiento de energía (grasas/aceites), constituyentes de membranas (glicerofosfolípidos, esfingolípidos, colesterol), hormonas (esteroides), vitaminas (liposolubles), portadores de oxígeno/ electrones (hemo), entre otros. Para los lípidos que son muy hidrofóbicos, como las grasas/aceites, el movimiento y almacenamiento en el ambiente acuoso del cuerpo requiere estructuras especiales. Otros lípidos anfipáticos, como los glicerofosfolípidos y los esfingolípidos se organizan espontáneamente en bicapas lipídicas cuando se colocan en agua. Curiosamente, las partes principales de muchos lípidos pueden derivarse de acetil-CoA.
Ácidos grasos
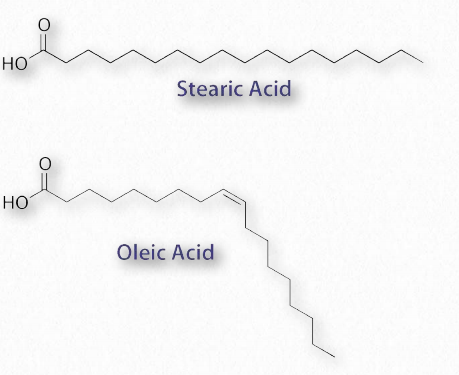
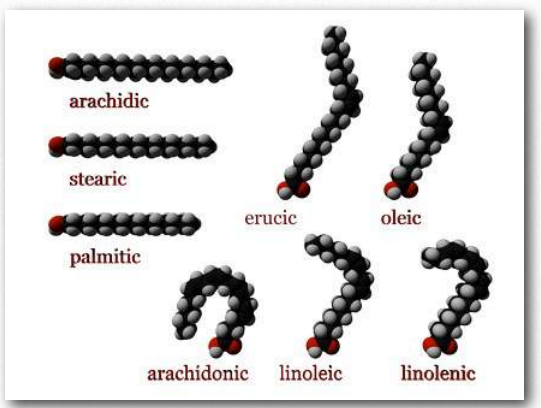
Los lípidos más ubicuos en las células son los ácidos grasos. Se encuentran en grasas, glicerofosfolípidos, esfingolípidos y que sirven como anclajes de membrana para proteínas y otras biomoléculas, los ácidos grasos son importantes para el almacenamiento de energía, estructura de membrana y como precursores de la mayoría de las clases de lípidos. Los ácidos grasos, como se puede observar en la Figura 2.190, se caracterizan por un grupo de cabeza polar y una larga cola de hidrocarburo. Los ácidos grasos con colas de hidrocarburos que carecen de dobles enlaces se describen como saturados, mientras que aquellos con uno o más dobles enlaces en sus colas se conocen como ácidos grasos insaturados. El efecto de los dobles enlaces en la cola de ácido graso es introducir una torcedura, o curva, en la cola, como se muestra para el ácido oleico.
El ácido esteárico, un ácido graso saturado, por el contrario, tiene una cola de hidrocarburo lineal. Las figuras 2.190-2.194 muestran los ácidos grasos saturados e insaturados más comunes. Los ácidos grasos con colas insaturadas tienen una temperatura de fusión más baja que aquellos con colas saturadas de la misma longitud. Las colas más cortas también disminuyen la temperatura de fusión. Estas propiedades se trasladan a las grasas/aceites que las contienen.
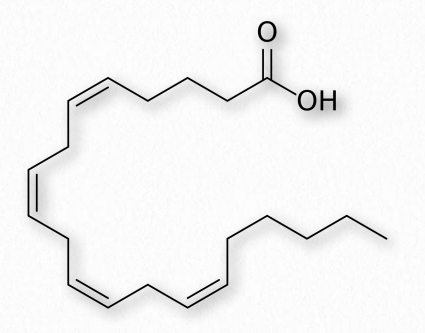
Los ácidos grasos con más de un doble enlace se denominan poliinsaturados. Las plantas son excelentes fuentes de ácidos grasos insaturados y poliinsaturados. La posición del doble enlace (s) en los ácidos grasos tiene consideraciones importantes tanto para su síntesis como para sus acciones en el organismo. Bioquímicamente, los dobles enlaces que se encuentran en los ácidos grasos se encuentran predominantemente en la configuración cis. Las llamadas grasas trans surgen como un subproducto químico de la hidrogenación parcial del aceite vegetal.
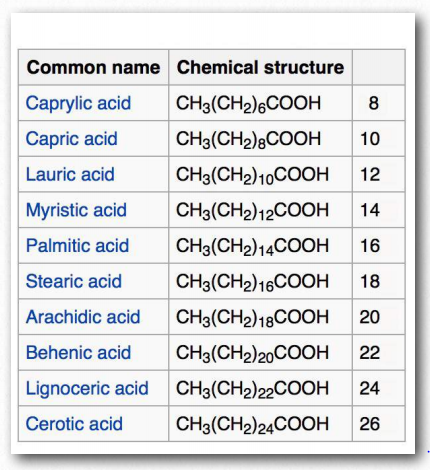
En humanos, el consumo de grasas trans eleva los niveles de lipoproteínas de baja densidad (LDL) y disminuye los niveles de lipoproteínas de alta densidad (HDL). Se cree que cada uno contribuye al riesgo de desarrollar enfermedad arterial coronaria. El más Figura 2.194 - Modelos de ácidos grasos. Los ácidos grasos comunes en nuestro cuerpo incluyen palmitato, estearato, oleato, linolenato, linolenato, linoleato y araquidonato. Dos ácidos grasos más cortos notables son el nonanoico (9 carbonos) y el ácido decanoico (10 carbonos), los cuales parecen tener efectos anticonvulsivos. El ácido decanoico inhibe directamente la neurotransmisión excitatoria en el cerebro y puede contribuir al efecto anticonvulsivo de la dieta cetogénica.
Numeración
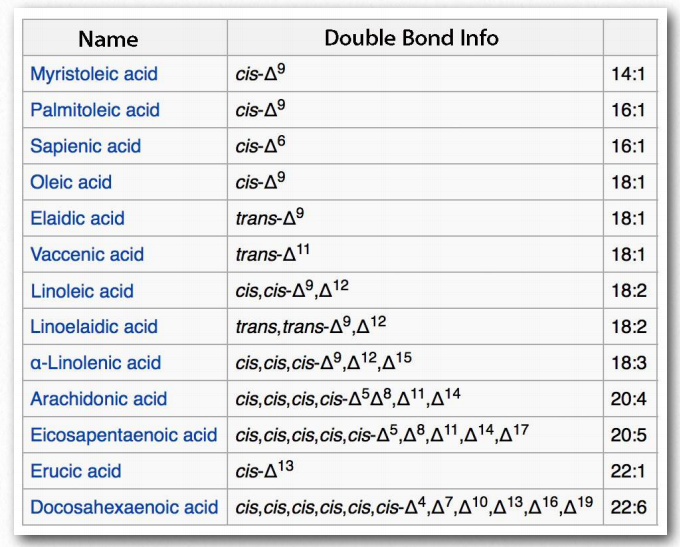
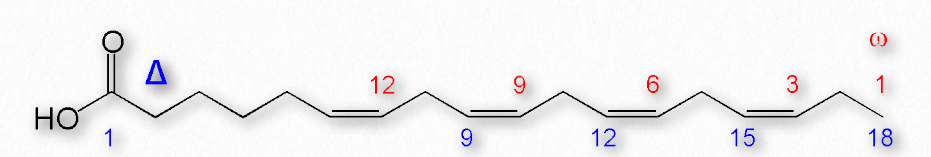
La Figura 2.195 muestra dos sistemas diferentes para localizar dobles enlaces en un ácido graso. El sistema ω cuenta los carbonos comenzando con el extremo metilo (mostrado en rojo) mientras que el sistema Δ cuenta desde el extremo carboxilo (mostrado en azul). Por ejemplo, un ácido graso ω-3 (omega 3) tendría un doble enlace en el tercer carbono desde el extremo metilo. En el sistema Δ, un ácido graso que tiene un doble enlace cis en el carbono 6, contando desde el extremo carboxilo, se escribiría como cis-δ6.
Los ácidos grasos se describen como ácidos grasos esenciales si deben estar en la dieta (no pueden ser sintetizados por el organismo) y ácidos grasos no esenciales si el organismo puede sintetizarlos. Los humanos y otros animales carecen de las enzimas desaturasas necesarias para hacer dobles enlaces en posiciones superiores a Δ-9, por lo que los ácidos grasos con dobles enlaces más allá de esta posición deben obtenerse en la dieta. El ácido linoleico y el ácido linolénico, ambos entran en esta categoría. Los ácidos grasos insaturados relacionados pueden elaborarse a partir de estos ácidos grasos, por lo que la presencia de ácidos linoleico y linolénico en la dieta elimina la necesidad de tener todos los ácidos grasos insaturados en la dieta. Tanto el ácido linoleico como el linolénico contienen 18 carbonos, pero el ácido linoleico es un ácido graso ω-6, mientras que el ácido linolénico es un ácido graso ω-3. Notablemente, los ácidos grasos ω-6 tienden a ser proinflamatorios, mientras que los ácidos grasos ω-3 son menores.

Grasas/aceites
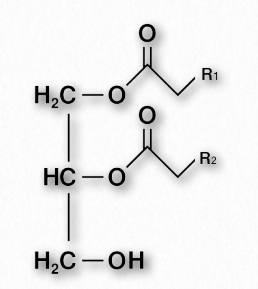
Las grasas y aceites son las formas primarias de almacenamiento de energía de los animales y también se conocen como triacilgliceroles y triglicéridos, ya que consisten en una molécula de glicerol unida a través de enlaces éster a tres ácidos grasos (Figura 2.196). Las grasas y aceites tienen la misma estructura básica. Damos el nombre grasa a aquellos compuestos que son sólidos a temperatura ambiente y el nombre aceite a los que son líquidos a temperatura ambiente. Tenga en cuenta que los aceites biológicos no son lo mismo que los aceites de petróleo.
Al aumentar el número de ácidos grasos insaturados (y la cantidad de insaturación en un ácido graso dado) en una grasa disminuye la temperatura de fusión de la misma. Organismos como el pescado, que viven en ambientes fríos, tienen grasas con más insaturación y es por ello que el aceite de pescado contiene ácidos grasos poliinsaturados.
Adipocitos
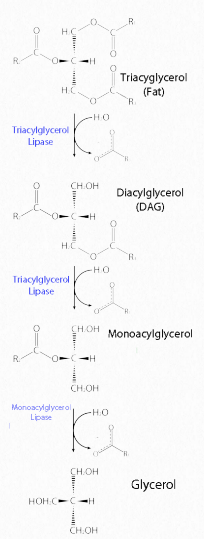
Las grasas se almacenan en el cuerpo en células especializadas conocidas como adipocitos. Las enzimas conocidas como lipasas liberan ácidos grasos de las grasas por reacciones de hidrólisis (Figura 2.197). La triacilglicercol lipasa (pancreática - Figura 2.198) es capaz de escindir los dos primeros ácidos grasos de la grasa. Una segunda enzima, monoacilglicerol lipasa, escinde el último ácido graso. Las grasas se pueden sintetizar reemplazando el fosfato en el ácido fosfatídico por un ácido graso.
Glicerofosfolipídos
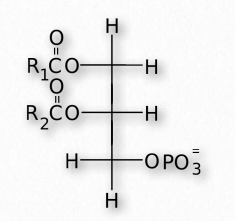
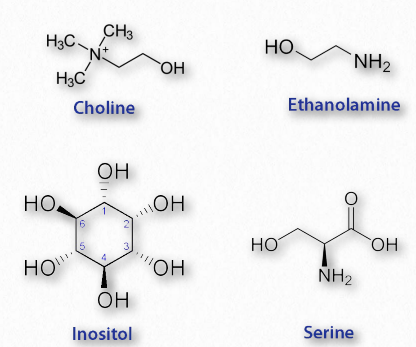
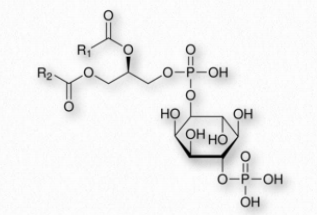
Los glicerofosfolípidos (fosfoglicéridos) son componentes importantes de la bicapa lipídica de las membranas celulares. Los fosfoglicéridos están estructuralmente relacionados con las grasas, ya que ambos se derivan del ácido fosfatídico (Figura 2.199). El ácido fosfatídico es un glicerofosfolípido simple que generalmente se convierte en compuestos de fosfatidilo. Estos se elaboran esterificando diversos grupos, como etanolamina, serina, colina, inositol, y otros (Figura 2.200) al fosfato de ácido fosfatídico. Todos estos compuestos forman bicapas lipídicas en solución acuosa, debido a su naturaleza anfifílica.
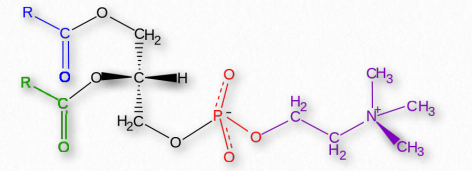
Fosfatidiletanolaminas
Dado que todos los glicerolípidos pueden tener una variedad de ácidos grasos en las posiciones 1 y 2 del glicerol, todos son familias de compuestos. Las fosfatidiletanolaminas se encuentran en todas las células vivas y son uno de los fosfátidos más comunes, constituyendo alrededor del 25% de ellas. Son constituyentes comunes del tejido cerebral y en la médula espinal, constituyendo hasta el 45% del total de fosfolípidos. Las fosfatidiletanolaminas se distribuyen asimétricamente a través de las membranas, ubicándose preferentemente en la valva interna (más cercana al citoplasma) de la membrana plasmática. Metabólicamente, las fosfatidiletanolaminas son precursores de fosfatidilcolinas. Fosfatidilserinas Las fosfatidilserinas son otro grupo de compuestos de fosfatidilo que se distribuyen preferentemente a través de la bicapa lipídica de la membrana plasmática. Al igual que las fosfatidiletanolaminas, las fosfatidilserinas se localizan preferentemente en la valva interna de la membrana plasmática. Cuando ocurre la apoptosis (suicidio celular), se pierde la distribución preferencial y las fosfatidilserinas aparecen en la valva externa donde sirven como señal a los macrófagos para que se unan y destruyan la célula.
Fosfatidilcolinas
Las fosfatidilcolinas (Figura 2.201) son otro grupo de componentes importantes de la membrana. Suelen encontrarse con mayor frecuencia en la valva externa de la membrana plasmática. Nutricionalmente, los compuestos se obtienen fácilmente a partir de huevos y soja. Las fosfatidilcolinas se mueven a través de las membranas mediante la proteína de transferencia de fosfatidilcolina (PCTP). Esta proteína, que es sensible a los niveles de fosfatidilcolinas, actúa para estimular la actividad de una tioesterasa (rompe los enlaces tioéster, como los acil-CoAs) y activa los factores de transcripción PAX3.
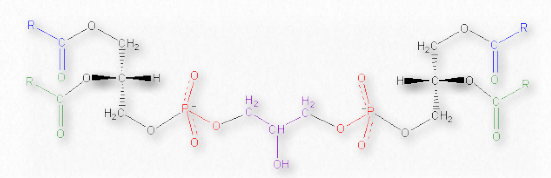
Cardiolipinas
Las cardiolipinas son un conjunto inusual de glicerofosfolípidos al contener dos cadenas principales de diacilglicerol unidas en el medio por un difosfoglicerol (Figura 2.202). Es un lípido de membrana importante, que constituye alrededor del 20% de la membrana mitocondrial interna y se encuentra en organismos desde bacterias hasta humanos. Tanto en plantas como en animales, se encuentra casi totalmente en la membrana mitocondrial interna.
Las moléculas parecen ser necesarias para que tanto el Complejo IV como el Complejo III de la cadena de transporte de electrones mantengan su estructura. La enzima ATP sintasa (Complejo V) del sistema de fosforilación oxidativa también se une a cuatro moléculas de cardiolipina. Se ha propuesto que la cardiolipina funciona como trampa de protones en el proceso de bombeo de protones por el Complejo IV.
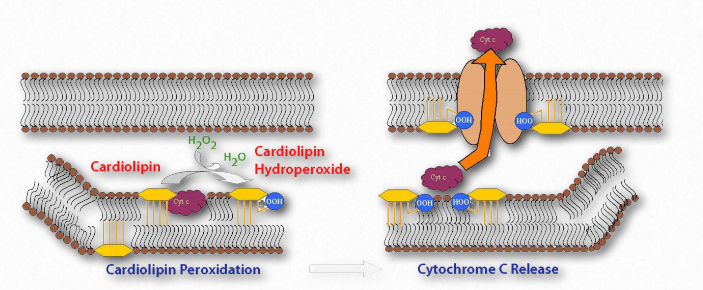
La cardiolipina también juega un papel en la apoptosis. Como se muestra en la Figura 2.203, la oxidación de cardiolipina por una oxigenasa específica de cardiolipina provoca que la cardiolipina se mueva de la membrana mitocondrial interna a la externa, ayudando a formar un poro permeable y facilitando el transporte del citocromo c fuera del espacio intermembrana y hacia el citoplasma - un paso en el proceso de apoptosis.
Diacilglicerol
El diacilglicerol (también llamado diglicérido y DAG - Figura 2.204) es un intermedio importante en las vías metabólicas. Se produce, por ejemplo, en la primera etapa de la hidrólisis de la grasa y también se produce cuando los lípidos de membrana, como PIP2 (fosfatidilinositol-4,5-bisfosfato) son hidrolizados por la fosfolipasa C en una cascada de señalización.
DAG es en sí mismo un compuesto de señalización, que se une a la proteína quinasa C para activarla para fosforilar sustratos. La síntesis de DAG comienza con glicerol-3-fosfato, que obtiene dos ácidos grasos de dos acil-CoAs para formar ácido fosfatídico. La desfosforilación del ácido fosfatídico produce DAG. El DAG también se puede refosforilar por la quinasa DAG para volver a hacer ácido fosfatídico u otro ácido graso se puede agregar para hacer grasa.
Inositol
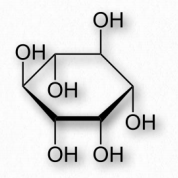
Aunque técnicamente no es un lípido en sí, el inositol se encuentra en muchos lípidos. El inositol es un derivado del ciclohexano que contiene seis grupos hidroxilo, uno en cada carbono (Figura 2.205. Tiene nueve estereoisómeros diferentes de los cuales uno, cis-1,2,3,5-trans-4,6-ciclohexanohexol (llamado mioinositol) es el más común. Tiene un sabor dulce (la mitad que la de la sacarosa).
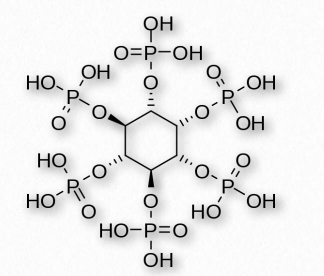
Existen numerosas formas fosforiladas del compuesto, desde un solo fosfato hasta seis (una en cada carbono). El ácido fítico, por ejemplo, en las plantas, tiene seis fosfatos (Figura 2.206) que utiliza para almacenar fosfato. El inositol se produce a partir de la glucosa y alguna vez se consideró vitamina B8, pero es elaborado por el organismo en cantidades adecuadas, por lo que ahora no se considera una vitamina. Las formas fosforiladas de inositol se encuentran en fosfoinosítidos, como PIP2 y PIP3, los cuales son importantes en los procesos de señalización. Algunos de estos incluyen señalización de insulina, catabolismo de grasa, regulación del calcio y ensamblaje del citoesqueleto.
Fosfoinosítidos
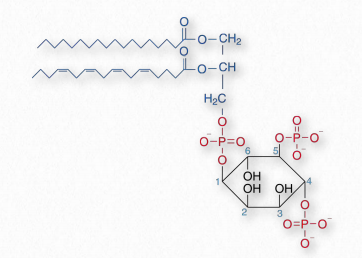
Los compuestos basados en fosfatidilinositol (PI) a menudo se denominan fosfoinosítidos. Estos compuestos tienen un papel importante en la señalización y el tráfico de membranas. Los hidroxilos en los carbonos 3,4 y 5 del anillo de inositol son dianas para la fosforilación por una variedad de quinasas. Se utilizan siete combinaciones diferentes. El impedimento estérico inhibe la fosforilación de los carbonos 2 o 6. El nombre de estos compuestos fosforilados sigue generalmente como PI (#P) P, PI (#P, #P) P o PI (#P, #P, #P) P donde #P se refiere al número del carbono donde se encuentra un fosfato. Por ejemplo, PI (3) P se refiere a un compuesto de fosfatidilo con un fosfato agregado a los carbonos 3 del anillo de inositol, mientras que PI (3,4,5) P es un compuesto de fosfatidilo con un fosfato agregado a los carbonos 3,4, y 5.
Fosfatidilinositol-4,5- bisfosfato
El fosfatidilinositol-4,5-bisfosfato (PIP2 - Figura 2.207) es un fosfolípido de membranas plasmáticas que funciona en la cascada de señalización de fosfolipasa C. En esta vía de señalización, la hidrólisis catalizada por la fosfolipasa C libera inositol-1,4,5- trisfosfato (IP3) y diacilglicerol. La síntesis de PIP2 comienza con fosfatidilinositol, el cual es fosforilado en la posición 4 seguido de fosforilación en la posición 5 por quinasas específicas.
PIP2 puede fosforilarse para formar la molécula de señalización conocida como fosfatidilinositol (3,4,5) -trifosfato (PIP3). Junto con PIP3, PIP2 sirve como fosfolípido de acoplamiento para el reclutamiento de proteínas que desempeñan un papel en cascadas de señalización. La unión de PIP2 también es requerida por los canales de potasio dirigidos hacia adentro.
Fosfatidilinositol (3,4,5) - trifosfato
El fosfatidilinositol (3,4,5) -trifosfato (PIP3) es una molécula importante para la activación de proteínas de señalización, como AKT, que activa las vías de señalización anabólicas relacionadas con el crecimiento y la supervivencia. PIP3 puede desfosforilarse por fosfatasa PTEN para producir PIP2 y puede sintetizarse a partir de PIP2 por acción quinasa de PI 3-quinasas Clase I. La actividad quinasa para sintetizar PIP3 resulta en el movimiento de las proteínas de unión a PIP3 a la membrana plasmática. Incluyen Akt/PKB, PDK1, Btk1 y ARNO y cada uno se activa mediante la unión a PIP3.
Plasmalógenos
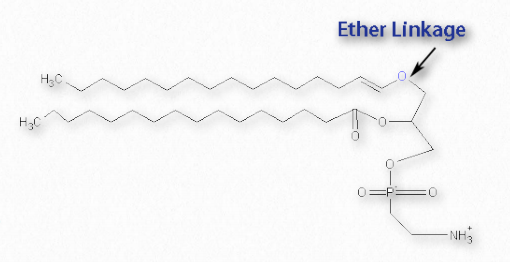
Una clase especial de los glicerofosfolípidos son los plasmalogenos (Figura 2.209). Difieren en contener un enlace vinil éter en la posición 1 del glicerol, en contraste con otros glicerofosfolípidos, que tienen un enlace éster en esta posición. La posición 2 de cada uno es un éster. El precursor para el enlace éter es típicamente un alcohol saturado de 16 ó 18 carbonos o un alcohol insaturado de 18 carbonos.
En la cola de fosfato, los grupos más comúnmente unidos son etanolamina o colina. Los plasmalógenos se encuentran abundantemente en humanos en el corazón (30-40% de los fosfolípidos de colina). El 30% de los glicerofosfolípidos en el cerebro son plasmalógenos y el 70% de los lípidos de etanolamina de la vaina de mielina de las células nerviosas son plasmalógenos.
Aunque no se entiende su función, se cree que los plasmalógenos pueden proporcionar cierta protección contra especies reactivas de oxígeno y tener papeles en la señalización.
Lecitina
La lecitina es un término genérico para una combinación de sustancias lipídicas que incluye ácido fosfórico, glicerol, glicolípidos, triglicéridos y fosfolípidos. La lecitina es un agente humectante útil con la emulsificación y encapsulación e incluso se usa como aditivo antilodo en lubricantes para motores. La lecitina se utiliza en barras de caramelo para evitar que el cacao y la manteca de cacao se separen Aunque se considera seguro como ingrediente alimentario, la lecitina puede ser convertida por las bacterias intestinales en trimetilamina-N-óxido que puede contribuir a la deposición de colesterol y a la aterosclerosis.
Esfingolípidos
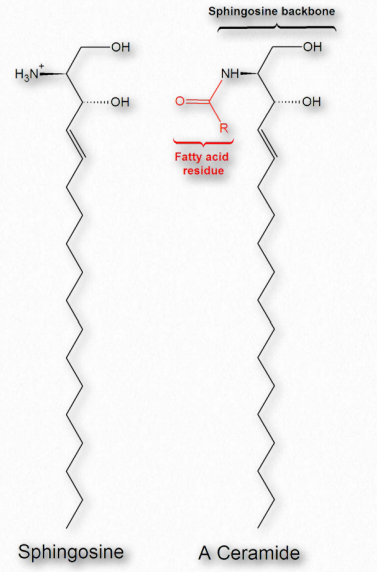
Los ácidos grasos también son componentes de una amplia clase de moléculas llamadas esfingolípidos. Los esfingolípidos son estructuralmente similares a los glicerofosfolípidos, aunque se sintetizan de forma completamente independiente a partir del ácido palmítico y el aminoácido serina. Los esfingolípidos reciben el nombre del aminoalcohol conocido como esfingosina (Figura 2.210), aunque no se sintetizan directamente a partir de él. La Figura 2.211 muestra la estructura generalizada de los esfingolípidos.
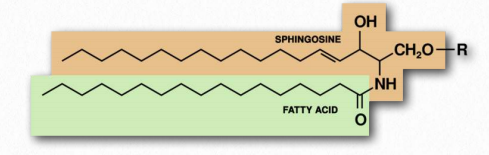
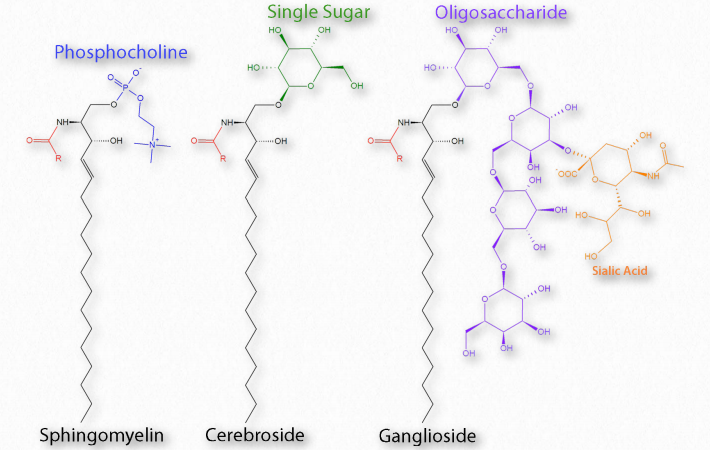
Si el grupo R es un hidrógeno, la molécula se llama ceramida. Cuando el grupo R es fosfoetanolamina, la molécula resultante es la esfingomielina, un componente importante de la vaina de mielina y las membranas lipídicas. Si en su lugar se agrega un solo azúcar simple, se crea un cerebrosido (Figura 2.212). La adición de un oligosacárido complejo crea un gangliósido.
Los esfingolípidos complejos pueden desempeñar un papel en el reconocimiento celular y la señalización. Los esfingolípidos se encuentran más abundantemente en la membrana plasmática y están casi completamente ausentes de las membranas mitocondriales y endoplásmicas del retículo. En los animales, los esfingolípidos en la dieta se han relacionado con una reducción del cáncer de colon, reducciones en los LDL y aumentos en HDL. Al igual que los glicerofosfolípidos, los esfingolípidos son anfifílicos. La mayoría de los esfingolípidos, excepto la esfingomielina, no contienen fosfato.
Eicosanoides
Los ácidos grasos elaborados a partir de ácidos grasos omega-6 y omega-3 incluyen tres ácidos grasos importantes que contienen 20 carbonos. Incluyen ácido araquidónico (un ácido graso ω-6 con cuatro dobles enlaces (Δ-5,8,11,14) - Figura 2.213), ácido eicosapentaenoico (un ácido graso ω-3 con cinco dobles enlaces, y ácido dihomo-γ-linolénico (un ácido graso ω-6 con tres dobles enlaces). La clase de compuestos conocidos como eicosanoides se elabora por oxidación de estos compuestos. Las subclases incluyen prostaglandinas, prostaciclinas, tromboxanos, lipoxinas, leucotrienos y endocannabinoides (Figuras 2.214-2.219). Los eicosanoides juegan papeles importantes que afectan la inflamación, la inmunidad, el estado de ánimo y el comportamiento.
Prostaglandinas
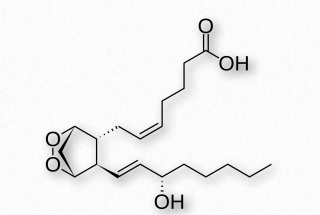
Una colección de moléculas que actúan como hormonas, las prostaglandinas se derivan del ácido araquidónico y tienen muchos efectos fisiológicos diferentes (incluso conflictivos). Estos incluyen constricción o dilatación de las células del músculo liso vascular, inducción del trabajo de parto, regulación de la inflamación y acción sobre el centro termorregulador del hipotálamo para inducir fiebre, entre otros.
Las prostaglandinas se agrupan con los tromboxanos (abajo) y prostaciclinas (abajo), como prostanoides. Los prostanoides, que contienen 20 carbonos, son una subclase de los eicosanoides. Las prostaglandinas se encuentran en la mayoría de los tejidos de organismos superiores. Son compuestos autocrinos o paracrinos producidos a partir de ácidos grasos esenciales. El precursor primario de las prostaglandinas es el ácido graso conocido como ácido araquidónico y la prostaglandina elaborada a partir del mismo se conoce como PGH2 (Figura 2.214), que, a su vez, es precursor de otras prostaglandinas, así como de las prostaciclinas y tromboxanos.
Interesantes prostaglandinas
PGD 2 - inhibe el crecimiento del folículo piloso, vasodilatador, causa constricción bronquial, mayor en pulmones de asmáticos que otros.
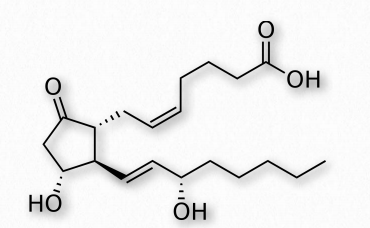
PGE 2 (Figura 2.215) - ejerce efectos en el trabajo de parto (ablandar el cuello uterino, contracción uterina), estimula la resorción ósea por los osteoclastos, induce fiebre, suprime la señalización del receptor de células T, vasodilatador, inhibe la liberación de noradrenalina desde las terminales nerviosas simpáticas. Es un potente activador de la vía de señalización Wnt.
Una prostaglandina puede tener efectos opuestos, dependiendo del receptor al que se une. La unión de PGE2 al receptor EP1 causa broncoconstricción y contracción del músculo liso, mientras que la unión de la misma molécula al receptor EP2 causa broncodilatación y relajación del músculo liso.
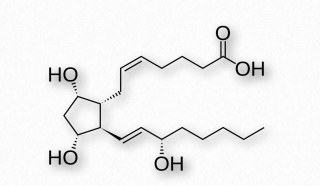
PGF 2α (Figura 2.216) - contracciones uterinas, induce parto, broncoconstricción.
IGP 2 - vasodilatación, broncodilatación, inhibición de la agregación plaquetaria.
Tromboxanes
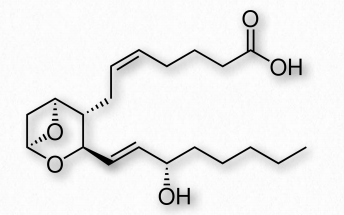
Los tromboxanos desempeñan un papel en la formación de coágulos y se llaman así por su papel en la trombosis. Son potentes vasoconstrictores y facilitan la agregación plaquetaria. También se sintetizan en plaquetas. Los efectos anticagulantes de la aspirina tienen sus raíces en la inhibición de la síntesis de PGH2, que es el precursor de los tromboxanos. Los tromboxanos más comunes son A2 (Figura 2.217) y B2.
Prostaciclina
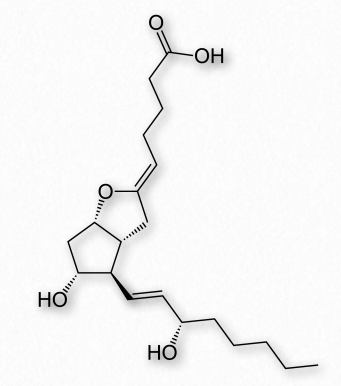
La prostaciclina (también conocida como prostaglandina I2 o PGI2 - Figura 2.218) contrarresta los efectos de los tromboxanos, inhibiendo la activación plaquetaria y actuando como vasodilatadores. Se produce a partir de PGH2 por acción de la enzima prostaciclina sintasa.
Leucotrienos
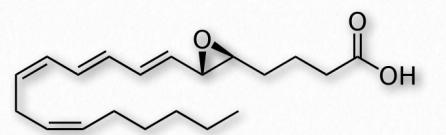
Otro grupo de compuestos eicosanoides son los leucotrienos (Figura 2.219). Al igual que las prostaglandinas, los leucotrienos están hechos de ácido araquidónico. La enzima que cataliza su formación es una dioxigenasa conocida como araquidonato 5-lipoxigenasa. Los leucotrienos están involucrados en la regulación de las respuestas inmunitarias. Se encuentran en leucocitos y otras células inmunocompetentes, como neutrófilos, monocitos, mastocitos, eosinófilos y basófilos. Los leucotrienos están asociados con la producción de histaminas y prostaglandinas, las cuales actúan como mediadores de la inflamación. Los leucotrienos también desencadenan contracciones en los músculos lisos de los bronquiolos. Cuando se sobreproducen, pueden desempeñar un papel en el asma y las reacciones alérgicas. Algunos tratamientos para el asma tienen como objetivo inhibir la producción o acción de leucotrienos.
Colesterol
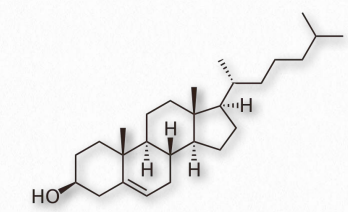
Podría decirse que ninguna biomolécula ha generado tanta discusión e interés como lo ha hecho el colesterol (Figura 2.220). Ciertamente, desde la perspectiva del comité del Premio Nobel, ni siquiera se acerca una molécula pequeña, ya que 13 personas han sido premiadas por trabajar en ella. La evidencia de la importancia del colesterol proviene del estudio del tejido cerebral donde comprende 10-15% de la masa seca.
Flexibilidad de membrana
En las células animales, el colesterol proporciona flexibilidad de membrana que permite el movimiento celular que está en contraste con las células vegetales y bacterianas con estructuras fijas. El colesterol se produce en muchas células del cuerpo, siendo el hígado la mayor cantidad. La vía anabólica que conduce a la síntesis del colesterol se conoce como la vía isoprenoide y las ramas de la misma conducen a otras moléculas incluyendo otras vitaminas liposolubles.

El colesterol solo se encuentra raramente en los procariotas (Mycoplasma, que lo requiere para el crecimiento, es una excepción) y se encuentra solo en cantidades traza en las plantas. En cambio, las plantas sintetizan compuestos similares llamados fitoesteroles (Figura 2.221). En promedio, el cuerpo de un macho adulto de 150 libras produce aproximadamente 1 gramo de colesterol al día, con un contenido total de aproximadamente 35 gramos.
Empaque
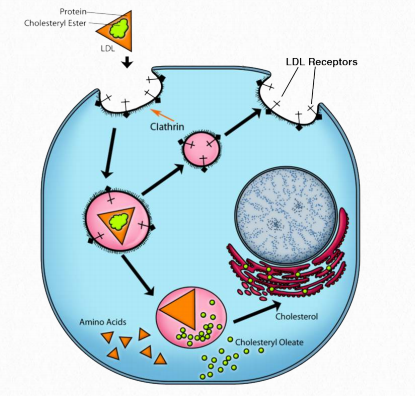
La hidrofobicidad del colesterol (y otros lípidos) requiere un empaquetado especial en complejos de lipoproteínas (llamados quilomicrones, VLDL, IDL, LDL y HDL) para el movimiento en el sistema linfático y el torrente sanguíneo. Aunque el colesterol puede ser producido por las células, también lo absorben del suministro de sangre al absorber LDLs que contienen colesterol directamente en un proceso llamado endocitosis mediada por receptores.
El daño oxidativo a los LDL puede llevar a la formación de placas ateroscleróticas y es por eso que el colesterol ha adquirido una imagen tan negativa en el ojo público. El hígado excreta el colesterol a través de la bilis para su eliminación en el sistema digestivo, pero el compuesto se recicla allí.
Reducción de los niveles de colesterol
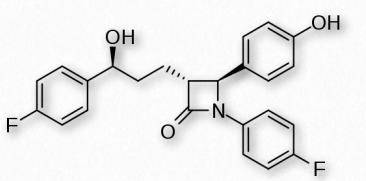
Las estrategias para reducir el colesterol en el cuerpo se enfocan principalmente en tres áreas: reducir el consumo, reducir la síntesis endógena y reducir el reciclaje. Consideraciones dietéticas, como el consumo de grasas saturadas versus grasas insaturadas, se debaten actualmente. Sin embargo, las grasas trans en la dieta se correlacionan con la incidencia de enfermedad coronaria. El consumo de verduras puede brindar alguna ayuda para reducir los niveles de colesterol reciclado en el sistema digestivo, debido a que los fitoesteroles vegetales compiten con el colesterol por la reabsorción y cuando esto sucede, un mayor porcentaje de colesterol sale del cuerpo en las heces. Los medicamentos relacionados con la penicilina también se utilizan para inhibir el reciclaje del colesterol. Uno de ellos es ezetimiba, que se muestra en la Figura 2.224.
Los defectos genéticos en el sistema de movimiento del colesterol son una causa de la rara enfermedad conocida como hipercolesterolemia familiar en la que la sangre de los individuos afectados contiene niveles peligrosamente altos de LDL. Si no se trata, la enfermedad suele ser mortal en los primeros 10-20 años de vida. Si bien los LDL han recibido (y merecen) una mala fama, otro grupo de complejos de lipoproteínas conocidos como los HDL (complejos de lipoproteínas de alta densidad) se conocen como “colesterol bueno” porque sus niveles se correlacionan con la eliminación de desechos (incluido el colesterol) de las arterias y reducen la inflamación.
Función de membrana
En las membranas, el colesterol es importante como aislante para la transmisión de señales en el tejido nervioso y ayuda a manejar la fluidez de las membranas en un amplio rango de temperaturas. Apilado en la bicapa lipídica, el colesterol disminuye la fluidez de una membrana y su permeabilidad a compuestos neutros, así como protones e iones de sodio. El colesterol puede desempeñar un papel en la señalización al ayudar con la construcción de balsas lipídicas dentro de la membrana celular.
Vitamina A
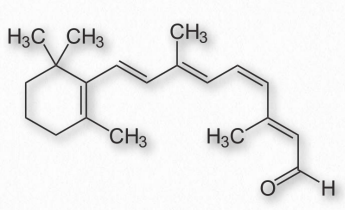
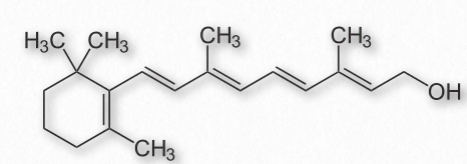
La vitamina A viene en tres formas químicas primarias, retinol (almacenamiento en hígado - Figura 2.225), retinal (papel en la visión - Figura 2.226) y ácido retinoico (roles en crecimiento y desarrollo). Todas las formas de vitamina A son diterpenoides y difieren solo en la forma química del grupo terminal. El retinol se usa principalmente como forma de almacenamiento de la vitamina.
El retinol se esterifica comúnmente a un ácido graso y se mantiene en el hígado. En niveles altos, el compuesto es tóxico. El retinol se obtiene en el organismo por hidrólisis del éster o por reducción de retinal. Es importante destacar que ni el retinal ni el retinol se pueden hacer a partir del ácido retinoico. El ácido retinoico es importante para la salud de la piel y los dientes, así como para el crecimiento óseo. Actúa en la diferenciación de células madre a través de un receptor celular específico de ácido retinoico.
Fuentes
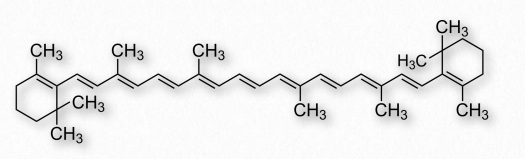
Buenas fuentes de vitamina A son el hígado y los huevos, así como muchas plantas, entre ellas las zanahorias, que tienen un precursor, el β-caroteno (Figura 2.227) a partir del cual se puede hacer retinol por acción de una dioxigenasa.
Sensibilidad a la luz El sistema de doble enlace conjugado en la cadena lateral de la vitamina A es sensible a la luz y puede cambiar entre las formas cis y trans al exponerse a ella. Es esta respuesta a la luz la que hace posible que la retina tenga un papel en la visión en los bastones y conos de los ojos. Aquí, la forma aldehído (retinal) se une a la proteína rodopsina en las membranas de las células bastonciformes y cónicas.
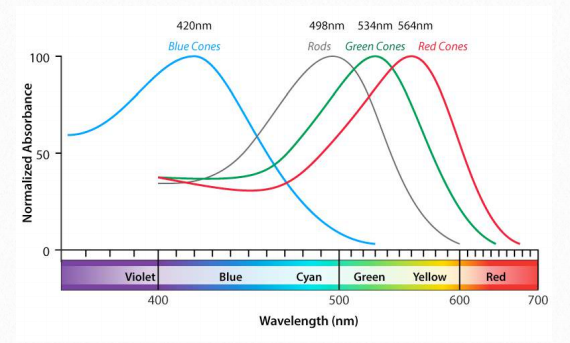
Cuando se expone a la luz de una longitud de onda particular, la “cola” de la molécula retiniana volteará hacia adelante y hacia atrás de cis a trans en el doble enlace en la posición 11 de la molécula. Cuando esto sucede, se genera una señal nerviosa que señala al cerebro de la exposición a la luz. Formas ligeramente diferentes de rodopsina tienen diferentes máximos de absorción que permiten que el cerebro perciba rojo, verde y azul específicamente y ensamblarlos en las imágenes que vemos (Figura 2.228). Los conos son las células responsables de la visión del color, mientras que los bastones están involucrados principalmente en la detección de luz en circunstancias de poca luz.
Deficiencia y superávit
La deficiencia de vitamina A es común en los países en desarrollo y fue inspiración para el diseño y síntesis del arroz dorado genéticamente modificado, que se utiliza como fuente de vitamina A para ayudar a prevenir la ceguera en los niños. La sobredosis de vitamina A, llamada hipervitaminosis A, es peligrosa y puede ser fatal. También se sospecha que el exceso de vitamina A está relacionado con la osteoporosis. En los fumadores, el exceso de vitamina A está vinculado a una mayor tasa de cáncer de pulmón, pero los no fumadores tienen una tasa reducida.
Vitamina D
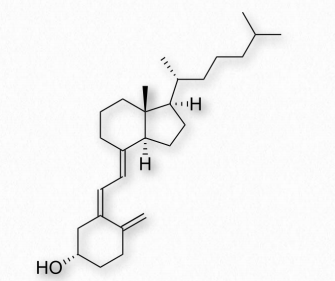
La forma activa de la vitamina D juega un papel importante en la absorción intestinal de calcio y fosfato y por lo tanto en huesos sanos. Técnicamente, la vitamina D ni siquiera es una vitamina, ya que es un compuesto hecho por el cuerpo. Más bien, se comporta más como una hormona.
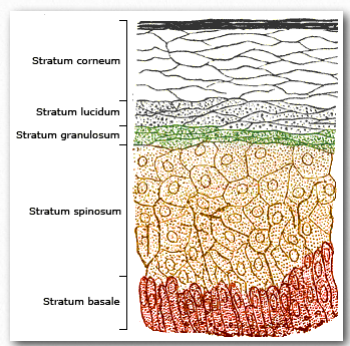
Derivada en última instancia del colesterol, la vitamina D puede elaborarse en una reacción catalizada por la luz ultravioleta. En la reacción, el intermedio 7-deshidrocolesterol se convierte en colecalciferol (vitamina D3) por la luz uv (Figura 2.229). La reacción ocurre más fácilmente en las dos capas inferiores de la piel que se muestran en la Figura 2.230.
Formas de vitamina D
Cinco compuestos diferentes son referidos como vitamina D. Son
Vitamina D1 - Una mezcla de ergocalciferol y lumisterol
Vitamina D2 - Ergocalciferol
Vitamina D3 - Colecalciferol Vitamina
D4 - 22-Dihidroergocalciferol Vitamina
D5 - Sitocciferol
La vitamina D3 es la forma más común utilizada en los suplementos vitamínicos y ésta y la vitamina D2 se obtienen comúnmente en la dieta, también. La forma activa de vitamina D, calcitriol (Figura 2.231), se elabora en el organismo en cantidades controladas. Esto procede a través de dos pasos a partir del colecalciferol. Primero, una hidroxilación en el hígado produce calcidiol y una segunda hidroxilación en el riñón produce calcitriol. Los macrófagos monocitarios también pueden sintetizar vitamina D y utilizan es como citocina para estimular el sistema inmune innato.
Mecanismo de acción
El calcitriol se mueve en el cuerpo unido a una proteína de unión a la vitamina D, que la entrega a los órganos diana. El calcitriol dentro de las células actúa uniéndose a un receptor de vitamina D (VDR), lo que resulta en la mayoría de los efectos fisiológicos de la vitamina. Después de unirse al calcitriol, el VDR migra al núcleo donde actúa como factor de transcripción para controlar los niveles de expresión de las proteínas transportadoras de calcio (por ejemplo) en el intestino. La mayoría de los tejidos responden al VDR unido al calcitriol y el resultado es la moderación de los niveles de calcio y fosfato en las células.
Deficiencia/exceso
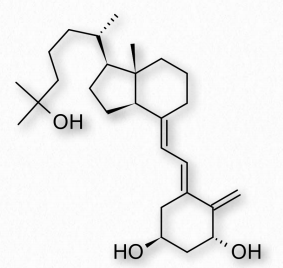
La deficiencia de vitamina D es una causa de la enfermedad conocida como raquitismo, que se caracteriza por huesos blandos y débiles y con mayor frecuencia se encuentra en niños. No es común en el mundo desarrollado, pero en otros lugares es cada vez más preocupante.
El exceso de vitamina D es raro, pero tiene efectos tóxicos, incluida la hipercalcemia, que resulta en depósitos dolorosos de calcio en los órganos principales. Los indicios de toxicidad por vitamina D son aumento de la micción y la sed. La toxicidad de la vitamina D puede provocar retraso mental y muchos otros problemas de salud graves.
Vitamina E
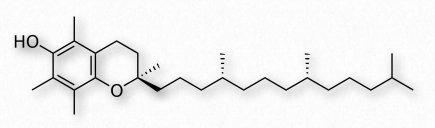
La vitamina E comprende un grupo de dos compuestos (tocoferoles y tocotrienoles - Figura 2.232) y estereoisómeros de cada uno. Se encuentra comúnmente en los aceites vegetales. Los compuestos actúan en las células como antioxidantes liposolubles. α-tocoferol (Figura 2.233), la forma más activa de la vitamina, actúa 1) a través del sistema protector de glutatión peroxidasa y 2) en membranas para interrumpir las reacciones en cadena de peroxidación lipídica. En ambas acciones, la vitamina E reduce los niveles de especies reactivas de oxígeno en las células.
Acción
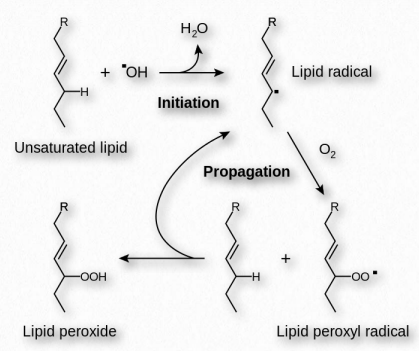
La vitamina E elimina los radicales de oxígeno (que poseen electrones desapareados) al reaccionar con ellos para producir un radical tocoferilo. Este radical de vitamina E puede ser convertido de nuevo a su forma original por un donante de hidrógeno. La vitamina C es uno de esos donantes. Actuando de esta manera, la Vitamina E ayuda a reducir la oxidación de compuestos fácilmente oxidados en las reacciones de peroxidación lipídica (Figura 2.234).
La vitamina E también puede afectar la actividad enzimática. El compuesto puede inhibir la acción de la proteína quinasa C en el músculo liso y activar simultáneamente la catálisis de la proteína fosfatasa 2A para eliminar los fosfatos, deteniendo el crecimiento del músculo liso.
Deficiencia/exceso
La deficiencia de vitamina E puede conducir a una mala conducción de las señales nerviosas y otros problemas derivados de problemas nerviosos. Los bajos niveles de la vitamina pueden ser un factor en los bajos pesos al nacer y los partos prematuros. La deficiencia, sin embargo, es rara, y no suele asociarse con la dieta.
El exceso de vitamina E reduce los niveles de vitamina K, reduciendo así la capacidad de coagular la sangre. La hipervitaminosis de la vitamina E junto con la aspirina puede poner en peligro la vida. En niveles más bajos, la vitamina E puede servir un papel preventivo con respecto a la aterosclerosis al reducir la oxidación de las LDL, un paso en la formación de placa.
Vitamina K
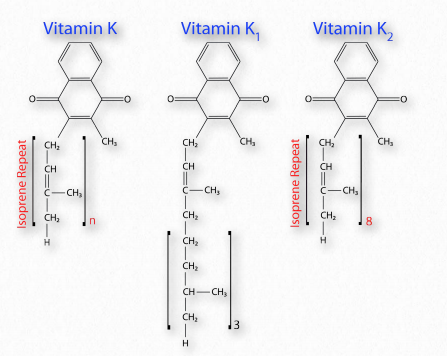
Al igual que las otras vitaminas liposolubles, la Vitamina K viene en múltiples formas (Figura 2.235) y se almacena en el tejido adiposo del cuerpo. Hay dos formas primarias de la vitamina - K1 y K2 y esta última tiene múltiples subformas. Las vitaminas K3, K4 y K5 se elaboran sintéticamente, no biológicamente.
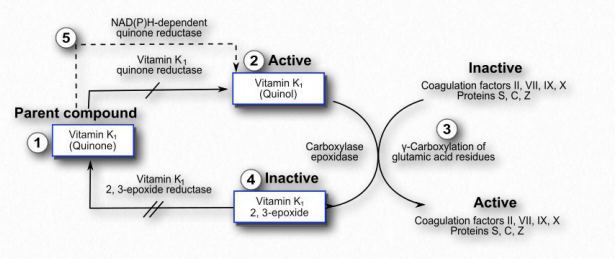
Acción
La vitamina K se utiliza como cofactor para enzimas que agregan grupos carboxilo a las cadenas laterales de glutamato de proteínas para aumentar su afinidad por el calcio. En los humanos se conocen dieciséis proteínas de este tipo. Incluyen proteínas involucradas en la coagulación de la sangre (protrombina (llamada Factor II), Factores VII, IX y X), metabolismo óseo (osteocalcina, también llamada proteína Gla ósea (BGP), proteína Gla de matriz (MGP) y periostina) y otras.
La modificación de la protrombina es un paso importante en el proceso de coagulación de la sangre (ver AQUÍ). Los niveles reducidos de vitamina K resultan en una menor coagulación de la sangre, un fenómeno que a veces se conoce como adelgazamiento de la sangre. Fármacos que bloquean el reciclaje de vitamina K (Figura 2.236) al inhibir la vitamina K epóxido reductasa, producen niveles más bajos de la vitamina y se emplean en tratamientos para personas propensas a la coagulación excesiva. La warfarina (cumadina) es uno de esos compuestos que actúa de esta manera y se usa terapéuticamente. Los individuos responden al fármaco de manera diferencial, requiriendo que periódicamente se les someta a pruebas para determinar los niveles de coagulación que poseen, para que no se produzca demasiado o muy poco.
Fuentes
La vitamina K1 es un estereoisómero del receptor de electrones del fotosistema I vegetal conocido como filoquinona y se encuentra abundantemente en vegetales de hoja verde. La filoquinona es una fuente de vitamina K, pero el compuesto se une fuertemente a las membranas tilacoides y tiende a tener baja biodisponibilidad. La vitamina K2 es producida por microbios en el intestino y es una fuente primaria de la vitamina. Los infantes en los primeros días antes de establecer su flora intestinal y las personas que toman antibióticos de amplio espectro pueden tener niveles reducidos, como resultado. La deficiencia dietética es poco frecuente en ausencia de daño al intestino delgado. Otros en riesgo de deficiencia incluyen personas con enfermedad renal crónica y cualquier persona que padezca una deficiencia de vitamina D. Las deficiencias producen síntomas de moretones fáciles, sangrado menstrual abundante, anemia y hemorragias nasales.
Esteroides
Los esteroides, como el colesterol, se encuentran en las membranas y actúan como hormonas de señalización al viajar por el cuerpo.
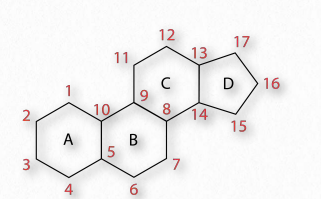
Todas las hormonas esteroides están hechas de colesterol y se agrupan en cinco categorías: mineralocorticoides (21 carbonos), glucocorticoides (21 carbonos), progestágenos (21 carbonos), andrógenos (19 carbonos) y estrógenos (18 carbonos).
Mineralocorticoides
Los mineralocorticoides son hormonas esteroides que influyen en los equilibrios de agua y electrolitos. La aldosterona (Figura 2.238) es la hormona mineralocorticoide primaria, aunque otras hormonas esteroides (incluida la progesterona) tienen algunas funciones como ésta. La aldosterona estimula a los riñones para reabsorber sodio, secretar potasio y reabsorber pasivamente el agua. Estas acciones tienen el efecto de aumentar la presión arterial y el volumen sanguíneo. Los mineralocorticoides son producidos por la zona glomerulosa de la corteza de la glándula suprarrenal.
Glucocorticoides
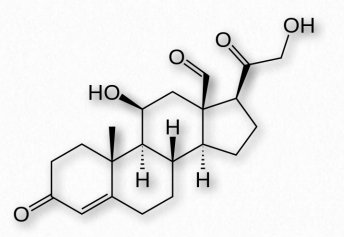
Los glucocorticoides (GC) se unen a los receptores de glucocorticoides que se encuentran en casi todas las células animales vertebradas y actúan en un mecanismo de retroalimentación en el sistema inmune para reducir su actividad. Los GC se utilizan para tratar enfermedades asociadas con sistemas inmunitarios hiperactivos. Estos incluyen alergias, asma y enfermedades autoinmunes- Figura 2.237 - Esquema de numeración de esteroides Imagen de Pehr Jacobson facilita. El cortisol (Figura 2.239) es un glucocorticoide importante con funciones cardiovasculares, metabólicas e inmunológicas. El glucocorticoide sintético conocido como dexametasona tiene aplicaciones médicas para tratar artritis reumatoide, broncoespasmos (en asma) e inflamación debido a su mayor potencia (25 veces) en comparación con el cortisol. Los glucocorticoides se producen principalmente en la zona fasciculata de la corteza suprarrenal.
Progestágenos
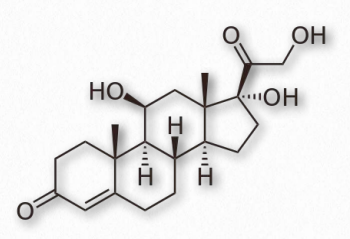
Los progestágenos (también llamados gestágenos) son hormonas esteroides que funcionan para activar el receptor de progesterona al unirse a él. Los progestágenos sintéticos se denominan progestinas. El progestágeno más común es la progesterona (también llamada P4 - Figura 2.240) y tiene funciones en el mantenimiento del embarazo. La progesterona es producida principalmente en la fase diestro del ciclo estral por el cuerpo lúteo de los ovarios de mamíferos. En el embarazo, la placenta asume la mayor parte de la producción de progesterona.
Andrógenos
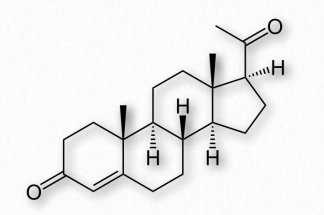
Los andrógenos son hormonas esteroides que actúan uniéndose a los receptores de andrógenos para estimular el desarrollo y mantenimiento de las características masculinas en vertebrados. Los andrógenos son precursores de estrógenos (ver más abajo). El andrógeno primario es la testosterona (Figura 2.241). Otros andrógenos importantes incluyen dihidrotestosterona (estimula la diferenciación del pene, escroto y próstata en el embrión) y androstenediona (precursor común de las hormonas masculinas y femeninas).
Estrógenos
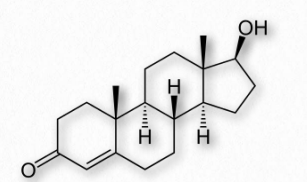
Las hormonas esteroideas estrógenas son una clase de compuestos con papeles importantes en los ciclos menstruales y estéticos. Son las hormonas sexuales femeninas más importantes. Los estrógenos actúan activando los receptores de estrógeno dentro de las células. Estos receptores, a su vez, afectan la expresión de muchos genes. Los estrógenos principales en las mujeres incluyen estrona (E1), estradiol (E2 - Figura 2.242) y estriol (E3). En los años reproductivos predomina el estradiol. Durante el embarazo predomina el estriol y durante la menopausia, la estrona es el estrógeno principal.
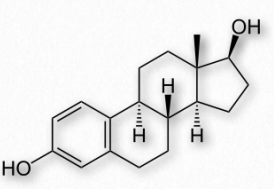
Los estrógenos se elaboran a partir de las hormonas andrógenas testosterona y androstenediona en una reacción catalizada por la enzima conocida como aromatasa. La inhibición de esta enzima con inhibidores de aromatasa, como el exemestano, es una estrategia para detener la producción de estrógenos. Esto puede ser parte de un tratamiento quimioterapéutico cuando están presentes tumores sensibles a estrógenos.
Cannabinoides
Los cannabinoides son un grupo de sustancias químicas que se unen y tienen efectos sobre los receptores cerebrales (receptores cannabinoides), reprimiendo la liberación de neurotransmisores. Las clases de estos compuestos incluyen endocannabinoides (hechos en el cuerpo), fitocannabinoides (elaborados en plantas, como la marihuana) y cannabinoides sintéticos (artificiales).
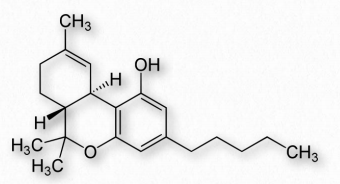
Los endocannabinoides son moléculas naturales derivadas del ácido araquidónico. Los receptores cannabinoides son muy abundantes, comprendiendo el mayor número de proteína G- 247 Figura 2.243 - Tetrahidrocannabinol - Ingrediente activo en receptores acoplados a marihuana que se encuentran en el cerebro. El fitocannabinoide más conocido es Δ-9- tetrahidrocannabinol (THC), el principal ingrediente psicoactivo (de los 85 cannabinoides) de la marihuana (Figura 2.243).
Anandamida
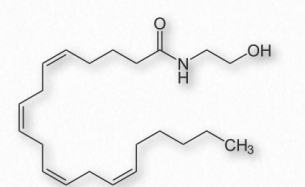
La anandamida (N-araquidonoiletanolamina - Figura 2.244) es un neurotransmisor endocannabinoide derivado del ácido araquidónico. Ejerce sus acciones principalmente a través de los receptores cannabinoides CB1 y CB2, los mismos unidos por el ingrediente activo de la marihuana, Δ9-tetrahidrocannabinol. La anandamida tiene un papel en estimular la comida/apetito y afectar la motivación y el placer. Se ha propuesto desempeñar un papel en el “corredor alto”, un efecto analgésico experimentado por el esfuerzo, especialmente entre los corredores. La anandamida parece afectar la función de la memoria en ratas.
La anandamida se ha encontrado en el chocolate y dos compuestos que imitan sus efectos (N-oleoiletanolamina y Nlinoleoiletanolamina) también están presentes. La enzima amida hidrolasa de ácidos grasos (FAAH) descompone la anandamida en ácido araquidónico libre y etanolamina.
Lipoxinas
Las lipoxinas (Figura 2.245) son compuestos eicosanoides implicados en la modulación de las respuestas inmunitarias y tienen efectos antiinflamatorios. Cuando las lipoxinas aparecen en la inflamación comienza el final del proceso. Las lipoxinas actúan para atraer a los macrófagos a las células apoptóticas en el sitio de la inflamación y son engullidos. Las lipoxinas actúan además para iniciar la fase de resolución del proceso de inflamación.
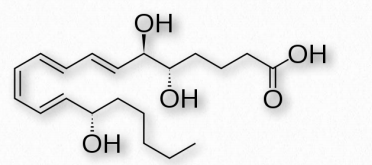
Al menos una lipoxina (LX4 activada por aspirina) tiene su síntesis estimulada por aspirina. Esto ocurre como un subproducto de la acetilación de la aspirina de COX-2. Cuando esto ocurre, la actividad catalítica de la enzima se redirige a la síntesis de ácido 15R-hidroxieicosatetraenoico (HETE) en lugar de prostaglandinas. El 15R-HETE es un procursor de lipoxinas de 15 epímeros, incluyendo LX4 desencadenadas por aspirina.
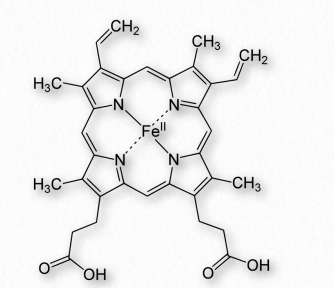
Heme
Los grupos hemo son una colección de cofactores proteínas/enzimas que contienen un gran anillo aromático heterocíclico conocido como anillo de porfirina con un ion ferroso (Fe++) en el medio. Un ejemplo de anillo de porfirina con un hierro (encontrado en Heme B de hemoglobina), se muestra en la Figura 2.246. Cuando están contenidas en una proteína, estas se conocen colectivamente como hemoproteínas (Figura 2.247).
El hemo, por supuesto, es un componente primario de la hemoglobina, pero también se encuentra en otras proteínas, como la mioglobina, citocromos, y las enzimas catalasa y succinato deshidrogenasa. Las hemoproteínas funcionan en el transporte de oxígeno, catálisis y transporte de electrones. El hemo se sintetiza en el hígado y la médula ósea en una vía que se conserva en una amplia gama de biología.

Porfobilinógeno
El porfobilinógeno (Figura 2.248) es una molécula de pirrol implicada en el metabolismo de la porfirina. Se produce a partir de aminolevulinato por acción de la enzima conocida como ALA deshidratasa. Porfobilinógeno es actuado por la enzima porfobilinógeno desaminasa. La deficiencia de esta última enzima (y otras en el metabolismo de la porfirina) puede dar como resultado una condición conocida como porfiria, que resulta en la acumulación de porfobilinógeno en el citoplasma de las células.
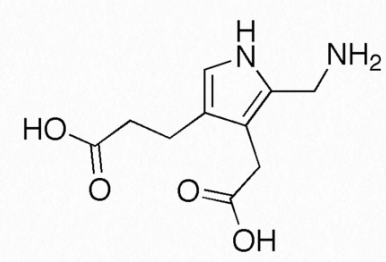
La enfermedad puede manifestarse con dolor abdominal agudo y numerosos problemas psiquiátricos. Se sospecha que tanto Vincent van Gogh como el rey `Jorge III sufrieron porfiria, lo que quizás provocó la “locura del rey Jorge III”. La porfiria también es considerada por algunos como el ímpetu para la leyenda de los vampiros que buscan sangre de las víctimas, ya que el color de la piel en formas no agudas de la enfermedad puede descolorarse, llevando a algunos a percibir eso como una deficiencia de hemoglobina y de ahí la “sed” de sangre imaginada para los vampiros.
Dolichols
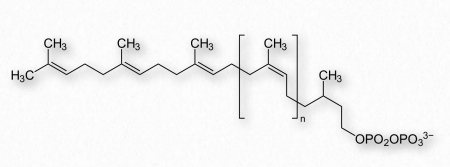
Dolichol es un nombre para un grupo de moléculas no polares hechas al combinar unidades de isopreno juntas. Las formas fosforiladas de dolicoles desempeñan un papel central en la N-glicosilación de proteínas. Este proceso, que ocurre en el retículo endoplásmico de células eucariotas, comienza con un pirofosfato de dolicol embebido en la membrana (Figura 2.249) al que se une un oligosacárido (ver también AQUÍ). Este oligosacárido contiene tres moléculas de glucosa, nueve moléculas de manosa y dos moléculas de N-acetilglucosamina.
Curiosamente, los azúcares se unen al pirofosfato de dolicol con el pirofosfato apuntando hacia afuera (lejos de) el retículo endoplásmico, pero después de la unión, el complejo de dolichol se voltea para que la porción de azúcar se sitúe en el interior del retículo endoplásmico. Allí, todo el complejo de azúcar se transfiere a la amida de una cadena lateral de asparagina de una proteína diana.
Las únicas cadenas laterales de asparagina a las que se puede hacer la unión son en proteínas donde ocurren las secuencias Asn-X-Ser o Asn-X-Thr. Los azúcares se pueden eliminar/agregar después de la transferencia a la proteína. Los niveles de dolicol en el cerebro humano aumentan con la edad, pero en las enfermedades neurodegenerativas, disminuyen.
Terpenos

Los terpenos son miembros de una clase de moléculas no polares hechas de unidades de isopreno. Los terpenos son producidos principalmente por las plantas y por algunos insectos. Los terpenoides son un grupo relacionado de moléculas que contienen grupos funcionales que carecen de terpenos.
Los terpenos tienen una variedad de funciones. En las plantas, a menudo juegan un papel defensivo protegiendo de los insectos. El nombre de terpeno proviene de la trementina, que tiene un olor como algunos de los terpenos. Los terpenos son componentes comunes de las resinas vegetales (piense en el pino) y son ampliamente utilizados en medicamentos y como fragancias. El lúpulo, por ejemplo, obtiene parte de su aroma y sabor distintivos a partir de los terpenos. No todos los terpenos, sin embargo tienen olor significativo.
Síntesis
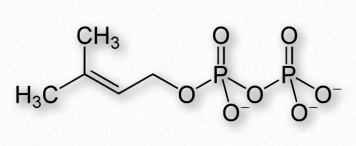
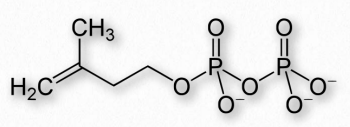
Los terpenos, al igual que los esteroides, se sintetizan comenzando con simples bloques de construcción conocidos como isoprenos. Hay dos de ellos: pirofosfato de dimetilalilo y el pirofosfato de isopentenilo relacionado y (Figuras 2.252 y 2.253) que combinan 1-2 unidades a la vez para hacer estructuras de orden superior. La síntesis de terpenos se superpone e incluye síntesis de esteroides.
Los terpenos y terpenoides se clasifican de acuerdo con la cantidad de unidades de isopreno que contienen. Incluyen hemiterpenos (una unidad), monoterpenos (dos unidades), sesquiterpenos (tres unidades), diterpenos (cuatro unidades), sesterterpenos (cinco unidades), triterpenos (seis unidades), sescuartos penos (siete unidades), tetraterrpenos (ocho unidades), politerpenos (muchas unidades). Otra clase de moléculas que contienen terpenos, los norisoterpenoides, surgen de reacciones catalizadas por peroxidasa en moléculas de terpeno.
Ejemplos
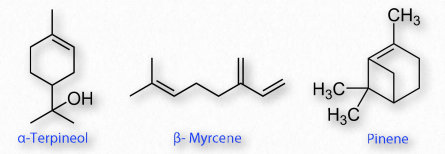
Los terpenos comunes incluyen monoterpenos de terpineol (lilas), limoneno (cítricos), mirceno (lúpulo), linalol (lavanda) y pineno (pino). Los terpenos de orden superior incluyen taxadieno (precursor diterpénico del taxol), licopeno (tetraterrenos), carotenos (tetraterrenos) y caucho natural (politerpenos).
Los precursores esteroideos pirofosfato de geranilo (derivado monoterpénico), pirofosfato de farnesil (derivado sesquiterpénico) y escualeno (triterpeno) son todos terpenos o derivados de ellos. La vitamina A y el fitol se derivan de los diterpenos.
Cafeína
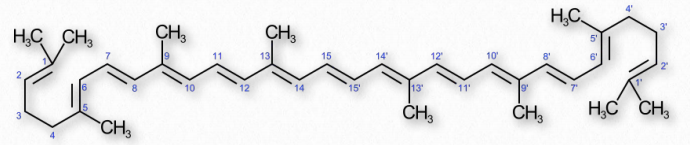
La cafeína es la droga psicoactiva más consumida en el mundo (Figura 2.255). Un alcaloide de metilxantina, la cafeína está estrechamente relacionada con la adenina y la guanina y esto es responsable de muchos efectos en el organismo. La cafeína bloquea la unión de la adenosina a su receptor y, en consecuencia, previene la aparición de somnolencia inducida por la adenosina. La cafeína cruza fácilmente la barrera hematoencefálica y estimula la liberación de neurotransmisores. La cafeína estimula porciones del sistema nervioso autónomo e inhibe la actividad de la fosfodiesterasa. Este último tiene el resultado de elevar los niveles de cAMP en las células, lo que activa la proteína quinasa A y activa la descomposición del glucógeno, inhibe la síntesis de TNF-α y leucotrienos, lo que resulta en la reducción de la inflamación y la inmunidad innata.
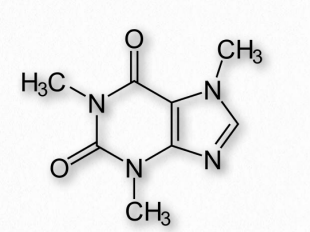
La cafeína también tiene efectos sobre el sistema colinérgico (inhibidor de la acetilcolinesterasa), es un antagonista del receptor de trifosfato de inositol 1 y es un activador independiente del voltaje de los receptores de riandina (un grupo de canales de calcio que se encuentran en el músculo esquelético, músculo liso y células del músculo cardíaco).
La vida media de la cafeína en el cuerpo varía considerablemente. En adultos sanos, tiene una vida media de aproximadamente 3-7 horas. La nicotina disminuye la vida media y los anticonceptivos y el embarazo pueden duplicarla. El hígado metaboliza la cafeína, por lo que la salud del hígado es un factor en la vida media. El CYP1A2 de la enzima oxidasa citocromo P450 es el principal responsable. La cafeína es un pesticida natural en las plantas, paralizando a los insectos depredadores.
Complejos de lipoproteínas y movimiento lipídico en el cuerpo
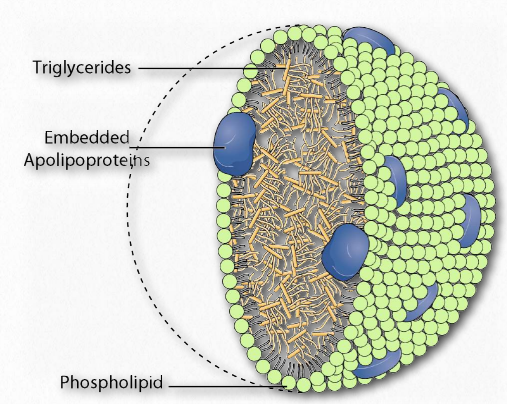
Los complejos de lipoproteínas son combinaciones de apolipoproteínas y lípidos unidos a ellos que solubilizan grasas y otras moléculas no polares, como el colesterol, para que puedan viajar en el torrente sanguíneo entre diversos tejidos del cuerpo. Las apolipoproteínas proporcionan la emulsificación necesaria para ello. Los complejos de lipoproteínas se forman en pequeñas “bolas” con las apolipoproteínas solubles en agua en el exterior y lípidos no polares, como grasas, ésteres de colesterilo y vitaminas solubles en grasa en el interior.
Se categorizan por sus densidades. Estas incluyen (de mayor densidad a la más baja) lipoproteínas de alta densidad (HDL), Lipoproteínas de Baja Densidad (LDL), Lipoproteínas de Densidad Intermedia (IDL), Lipoproteínas de Muy Baja Densidad (VLDL) y los quilomicrones. Estas partículas se sintetizan en el hígado y el intestino delgado.
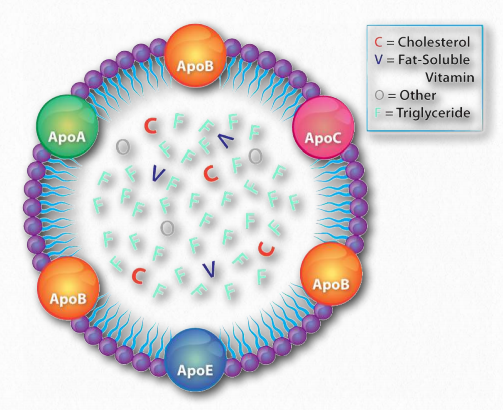
Apolipoproteínas
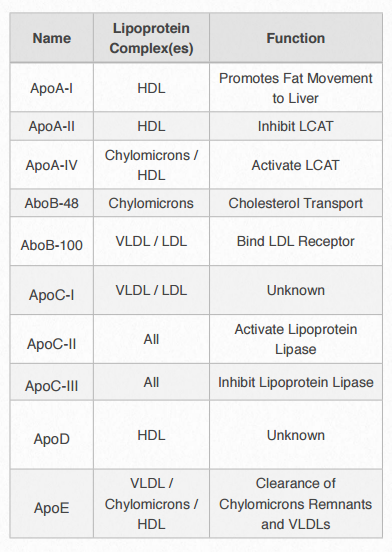
Cada complejo de lipoproteínas contiene un conjunto característico de apolipoproteínas, como se muestra en la Figura 2.256. ApoC-II y ApoC-III son notables por su presencia en todos los complejos de lipoproteínas y el papel que desempeñan en activar (apoC-II) o inactivar (apoC-III) lipoproteína lipasa. La lipoproteína lipasa es una enzima celular que cataliza la descomposición de la grasa de los complejos. ApoE (ver abajo) es útil para ayudar a predecir la probabilidad de ocurrencia de la enfermedad de Alzheimer en un paciente.
Edición genética
ApoB-48 y ApoB-100 son interesantes por ser codificados por el mismo gen, pero se produce un evento único de edición de secuencias de ARNm que convierte una en la otra. ApoB-100 se hace en el hígado, pero ApoB-48 se hace en el intestino delgado. El intestino delgado contiene una enzima que desamina la citidina en el nucleótido #2153 del ARNm común. Esto lo cambia a una uridina y cambia el codón en el que se encuentra de CAA (códigos para glutamina) a UAA (codón de parada). El hígado no contiene esta enzima y no realiza el cambio en el ARNm. En consecuencia, se sintetiza una proteína más corta en el intestino (apoB-48) que la que se elabora en el hígado (apoB-100) utilizando la misma secuencia génica en el ADN.
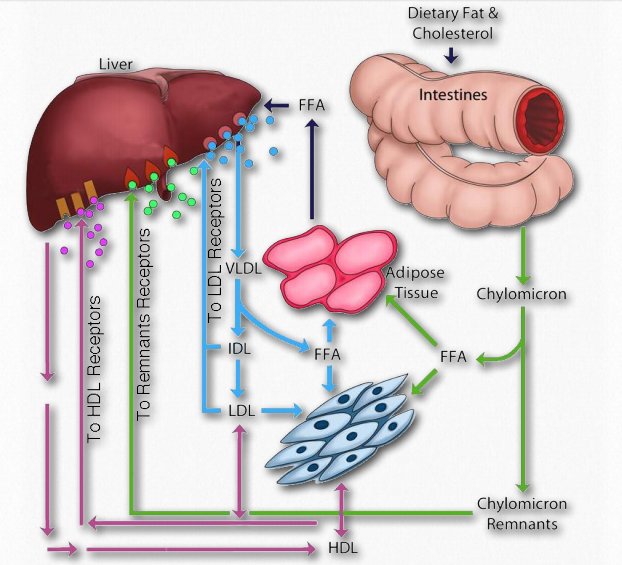
Movimiento
El movimiento de las grasas en el cuerpo es importante porque no se almacenan en todas las células. Sólo las células especializadas llamadas adipocitos almacenan grasa. Existen tres vías relevantes en el cuerpo para mover los lípidos. Como se describe a continuación, son 1) la vía exógena; 2) la vía endógena y 3) la vía de transporte inverso.
Vía exógena
La grasa dietética que ingresa al cuerpo desde el sistema intestinal debe transportarse, según corresponda, a lugares que la necesiten o almacenarla. Esta es la función de la vía exógena del movimiento lipídico en el cuerpo. Todos los lípidos de la dieta (grasas, colesterol, vitaminas liposolubles y otros lípidos) son movidos por él. En el caso de la grasa dietética, inicia su andadura después de la ingestión primero al ser solubilizada por los ácidos biliares en el tracto intestinal. Después de pasar por el estómago, las lipasas pancreáticas recortan dos ácidos grasos de la grasa, dejando un monoacil glicerol. Los ácidos grasos y el monoacil glicerol son absorbidos por las células intestinales (enterocitos) y se vuelven a ensamblar en una grasa, y luego ésta se mezcla con fosfolípidos, ésteres de colesterol y apolipoproteína B-48 y se procesa para formar quilomicrones (Figuras 2.258 y 2.259) en el aparato de Golgi y endoplásmico liso retículo.
Exocitosis
Éstos son exocitados de la célula a capilares linfáticos llamados lacteos. Los quilomicrones pasan a través de los lacteos y entran al torrente sanguíneo por la vena subclavia izquierda. Dentro del torrente sanguíneo, la lipoproteína lipasa descompone las grasas haciendo que el quilomicrón se encoja y se convierta en lo que se conoce como un remanente de quilomicrón. Conserva su colesterol y otras moléculas lipídicas.
Los restos de quilomicrón viajan al hígado donde son absorbidos (Figura 2.260). Esto se logra mediante receptores en el hígado que reconocen y se unen a la ApoE de los quilomicrones. Los complejos unidos son luego internalizados por endocitosis, degradados en los lisosomas, y el colesterol se desembolsa en las células hepáticas.
Vía endógena
El hígado juega un papel central en el manejo de las necesidades del cuerpo de lípidos. Cuando el cuerpo necesita lípidos o cuando la capacidad del hígado para contener más lípidos de los que suministra la dieta, el hígado empaqueta grasas y ésteres de colesterilo en complejos de lipoproteínas de muy baja densidad (VLDL) y los exporta por vía endógena. Los complejos de VLDL contienen apolipoproteínas ApoB-100, ApoC-I, ApoC-II, ApoC-III y ApoE. Las VLDL ingresan a la sangre y viajan a los músculos y al tejido adiposo donde la lipoproteína lipasa es activada por ApoC-II. En las células musculares, los ácidos grasos liberados son absorbidos y oxidados. Por el contrario, en los adipoctyes, los ácidos grasos se recogen y vuelven a ensamblar en triacilglicéridos (grasas) y se almacenan en gotitas de grasa. La eliminación de grasa de las VLDL hace que se encojan, primero a complejos de lipoproteínas de densidad intermedia (IDL) (también llamados restos de VLDL) y luego a complejos de lipoproteínas de baja densidad (LDL).
La reducción de las VLDL se acompaña de pérdida de apolipoproteínas de manera que las LDL están compuestas principalmente por ApoB-100. Este complejo lipoproteico es importante porque las células tienen receptores para que se una e internalice por endocitosis mediada por receptores (Figura 2.261). Hasta este punto, el colesterol y los ésteres de colesterilo han viajado en quilomicrones, VLDL e IDL ya que la grasa ha sido despojada. Para que los compuestos de colesterol ingresen a la célula a partir de los complejos de lipoproteínas, deben ser internalizados por las células y ese es el trabajo de la endocitosis mediada por receptores.
Vía de transporte inverso
Otra consideración importante del movimiento de los lípidos en el cuerpo es la vía de transporte inverso (Figura 2.260). También se le llama la vía de transporte inverso del colesterol, ya que el colesterol es la molécula primaria involucrada. Esta vía involucra la última clase de complejos de lipoproteínas conocidos como Lipoproteínas de Alta Densidad (HDL). A diferencia de los LDL, que comúnmente se conocen como “colesterol malo” (ver también a continuación), los HDL se conocen como “colesterol bueno”.
Los HDL se sintetizan en el hígado y el intestino delgado. Contienen poco o ningún lípido cuando se hacen (llamados HDL empobrecidos), pero cumplen el papel de “carroñeros” para el colesterol en la sangre y de restos de otros complejos de lipoproteínas (dañados) en la sangre. Para realizar su tarea, las HDL portan la enzima conocida como lecitincolesterol acil transferasa (LCAT), que utilizan para formar ésteres de colesterilo utilizando ácidos grasos de lecitina (fosfatidilcolina) y luego los internalizan.
El colesterol utilizado para este propósito proviene del torrente sanguíneo, de los macrófagos y de las células espumosas (complejos macrófago-LDL - Figura 2.262). La adición de ésteres de colesterilo provoca que el HDL se hinche y Figura 2.261 - El proceso de endocitosis mediada por receptores Imagen de Aleia Kim cuando está madura, devuelve su carga de colesterol de regreso al hígado o, alternativamente, a moléculas de LDL para endocitosis. Los HDL tienen el efecto de disminuir los niveles de colesterol y es por esa razón que se les describe como “colesterol bueno”.
Regulación de la transferencia de lípidos
Es importante que las células obtengan alimento cuando lo necesitan por lo que es crítico cierto control del movimiento de los nutrientes. El hígado, que juega el papel central en la modulación de los niveles de glucosa en sangre, también es importante para desempeñar el mismo papel para los lípidos. Realiza esta tarea el uso de receptores de LDL especializados en su superficie. Los receptores de LDL hepáticos se unen a las LDL que no fueron absorbidas por otras células en su camino a través del torrente sanguíneo. Los niveles altos de LDL son una señal al hígado para reducir la creación de VLDLs para su liberación.
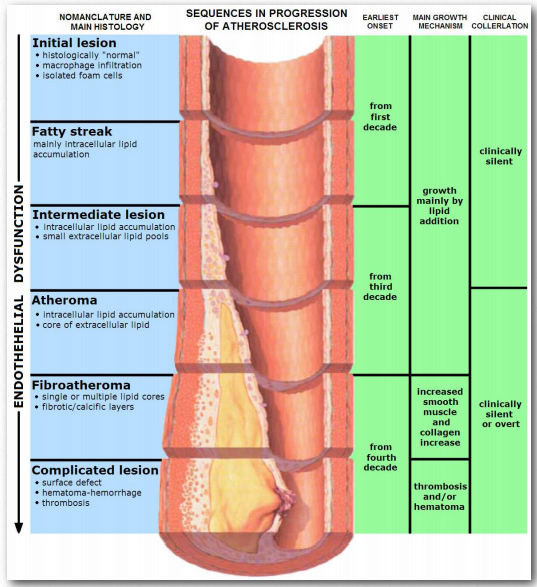
Las personas con la enfermedad genética conocida como hipercolesterolemia familiar, que se manifiesta con niveles peligrosamente altos de LDL, carecen de receptores de LDL que funcionen correctamente en sus células hepáticas.Figura 2.263 - Progresión de la aterosclerosis Wikipedia Figura 2.263 - Progresión de la aterosclerosis Wikipedia Figura 2.263 - Progresión de la aterosclerosis Wikipedia
En pacientes de esta enfermedad, el hígado nunca recibe la señal de que los niveles de LDL son altos. De hecho, para el hígado, parece que todos los VLDL y LDL están siendo absorbidos por los tejidos periféricos, por lo que crea más VLDL para intentar potenciar los niveles. Sin tratamiento, la enfermedad solía ser fatal temprano, pero los medicamentos más nuevos como las estatinas han aumentado significativamente la duración de la vida de los pacientes. Las necesidades celulares de los contenidos de LDL están directamente ligadas a los niveles de síntesis de los receptores de LDL en sus membranas. A medida que las células necesitan más colesterol, su síntesis de componentes para receptores sube y disminuye a medida que disminuye la necesidad.
Colesterol bueno/colesterol malo
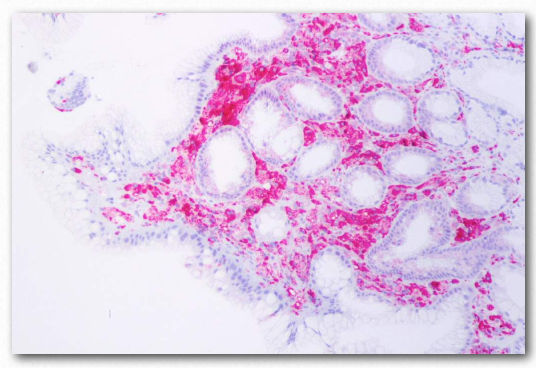
Se acepta comúnmente que los niveles de “colesterol alto” no son saludables. Esto se debe, al menos indirectamente, a los portadores primarios de colesterol, los LDL. Una función primaria de las LDL es administrar colesterol y otros lípidos directamente a las células mediante endocitosis mediada por receptores (Figura 2.237). Sin embargo, los niveles altos de LDL se correlacionan con la formación de placas ateroscleróticas (Figura 2.263 y 2.264) y la incidencia de aterosclerosis, lo que lleva a la descripción de las mismas como “colesterol malo”. Esto se debe a que cuando los niveles de LDL son muy altos, comienza la formación de placa. Se piensa que las especies reactivas de oxígeno (mayor en la sangre de los fumadores) provocan la oxidación parcial de los grupos ácidos grasos en los LDL. Cuando los niveles son altos, tienden a acumularse en la matriz extracelular de las células epiteliales en el interior de las arterias. Los macrófagos del sistema inmune absorben los LDL dañados (incluido el colesterol).
Dado que los macrófagos no pueden controlar la cantidad de colesterol que absorben, el colesterol comienza a acumularse en ellos y adquieren una apariencia que lleva a que sean descritas como “células espumosas”. Con demasiado colesterol, las células espumosas, sin embargo, están condenadas a morir por el proceso de muerte celular programada (apoptosis). La acumulación de estos, junto con el tejido cicatricial de la inflamación dan como resultado la formación de una placa. Las placas pueden crecer y bloquear el flujo de sangre o trozos de ellas pueden desprenderse y tapar aberturas más pequeñas en el suministro de sangre, lo que finalmente conduce a un ataque cardíaco o un derrame cerebral.
Colesterol bueno
Por otro lado, los niveles altos de HDL están inversamente correlacionados con la aterosclerosis y la enfermedad arterial. Los HDL empobrecidos son capaces de eliminar el colesterol de las células de espuma. Esto ocurre como resultado del contacto entre la proteína ApoA-I del HDL y una proteína de transporte en la célula espumosa (ABC-G1). Otra proteína de transporte en la célula espumosa, ABCA-1 transporta colesterol extra desde el interior de la célula a la membrana plasmática donde es absorbido hacia el HDL y devuelto al hígado o a los LDL por la vía de transporte inverso del colesterol.
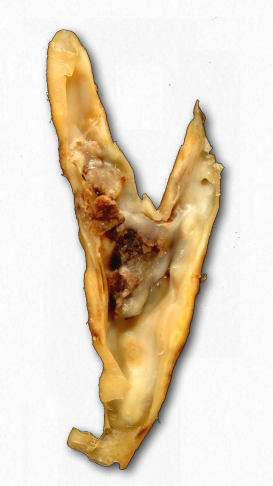
La deficiencia del gen ABCA-1 conduce a la enfermedad de Tánger. En esta condición, los HDL están casi totalmente ausentes porque permanecen vacíos como consecuencia de no poder absorber el colesterol de las células espumosas, por lo que son destruidos por el cuerpo.
ApoE y enfermedad de Alzheimer
La ApoE es un componente de los quilomicrones y también se encuentra en el cerebro, los macrófagos, los riñones y el bazo. En humanos, se encuentra en tres alelos diferentes, E2, E3 y E4. El alelo E4 (presente en aproximadamente 14% de la población) se asocia con una mayor probabilidad de contraer la enfermedad de Alzheimer. Las personas heterocigóticas para el alelo tienen 3 veces más probabilidades de contraer la enfermedad y las homocigóticas para ello tienen 15 veces más probabilidades de hacerlo. No se sabe por qué este gen o alelo está ligado a la enfermedad. Los tres alelos difieren solo ligeramente en la secuencia de aminoácidos, pero los cambios sí causan notables diferencias estructurales. El alelo E4 se asocia con un aumento de los niveles de iones calcio y apoptosis después de la lesión. La enfermedad de Alzheimer se asocia con la acumulación de agregados del péptido β- amiloide. ApoE mejora la descomposición proteolítica de la misma y la isoforma E4 no es tan eficiente en estas reacciones como las otras isoformas.