3.1: Conceptos Básicos en Membranas
- Page ID
- 53125

Fuente: BiochemFFA_3_1.pdf. Todo el libro de texto está disponible gratuitamente de los autores en http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy
Bicapas lipídicas
La membrana protectora alrededor de las células contiene muchos componentes, incluyendo colesterol, proteínas, glicolípidos, glicerofosfolípidos y esfingolípidos. Los dos últimos de estos, cuando se mezclan vigorosamente con agua, formarán espontáneamente lo que se denomina bicapa lipídica (Figura 3.1), que sirve como límite protector para la célula que es en gran parte impermeable al movimiento de la mayoría de los materiales a través de ella. Con las notables excepciones del agua, el dióxido de carbono, el monóxido de carbono y el oxígeno, la mayoría de los polares/iónicos requieren proteínas de transporte para ayudarlos a navegar eficientemente a través de la bicapa. El movimiento ordenado de estos compuestos es crítico para que la célula pueda 1) obtener alimentos para obtener energía; 2) exportar materiales; 3) mantener el equilibrio osmótico; 4) crear gradientes para el transporte secundario; 5) proporcionar fuerza electromotriz para la señalización nerviosa; y 6) almacenar energía en gradientes electroquímicos para ATP producción (fosforilación oxidativa o fotosíntesis). En algunos casos, se requiere energía para mover las sustancias (transporte activo).
Difusión facilitada
En otros casos, no se requiere energía externa y se mueven por difusión a través de canales celulares específicos. Esto se conoce como difusión facilitada. Antes de discutir el movimiento de materiales a través de las membranas, es apropiado discutir la composición de las membranas celulares. Las membranas plasmáticas difieren de las paredes celulares tanto en los materiales que las componen como en su flexibilidad. Las paredes celulares se cubrirán cerca del final de este capítulo.
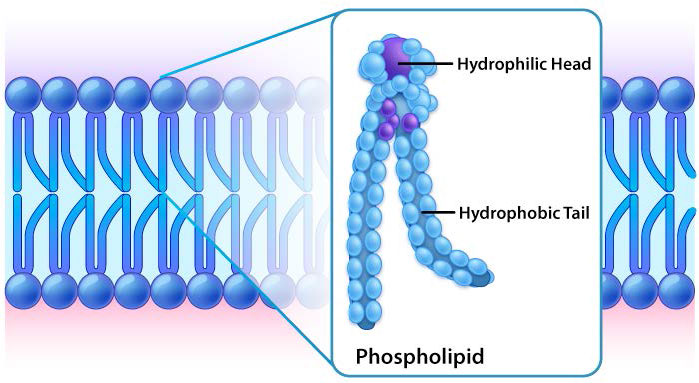
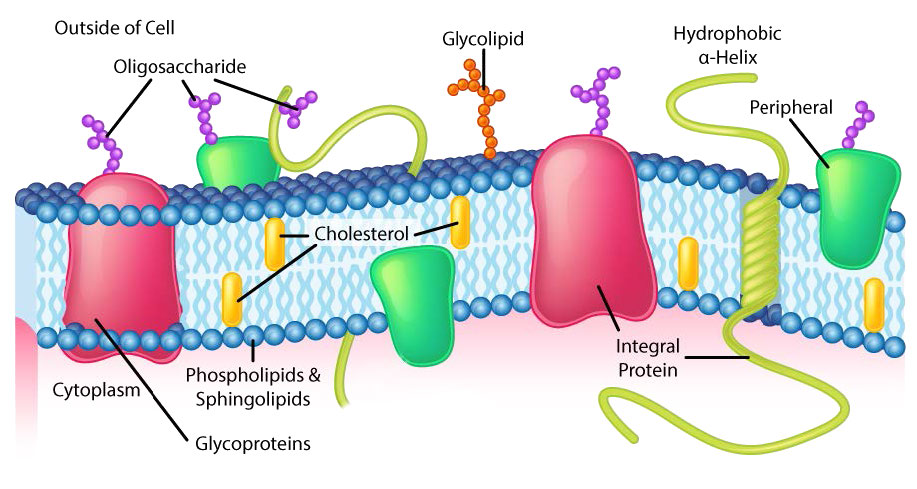
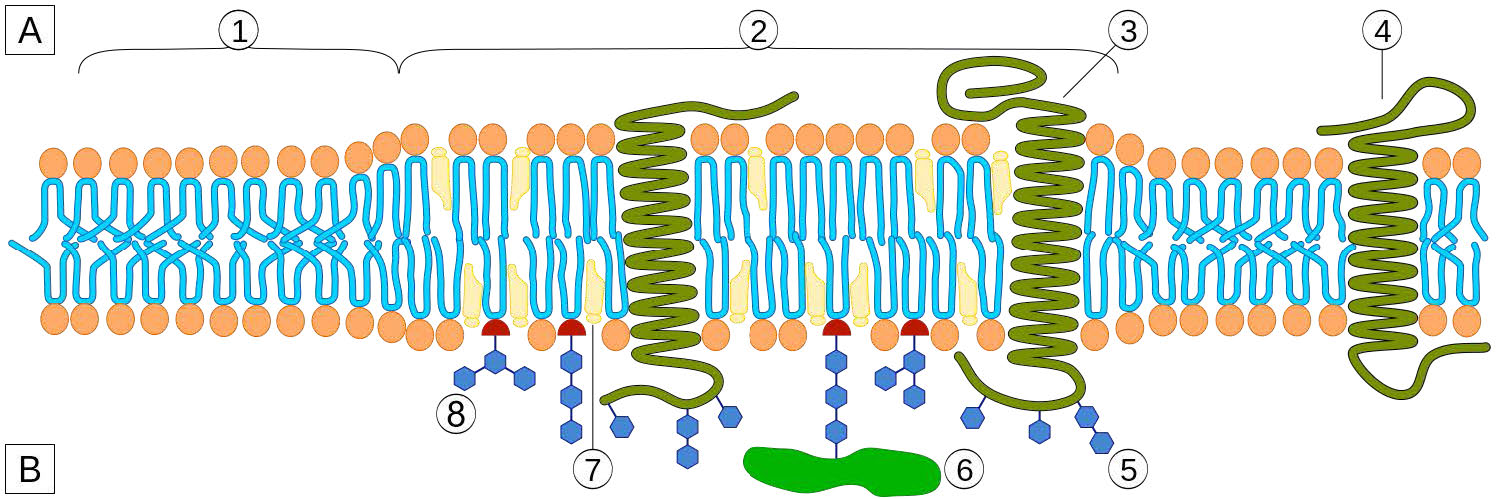
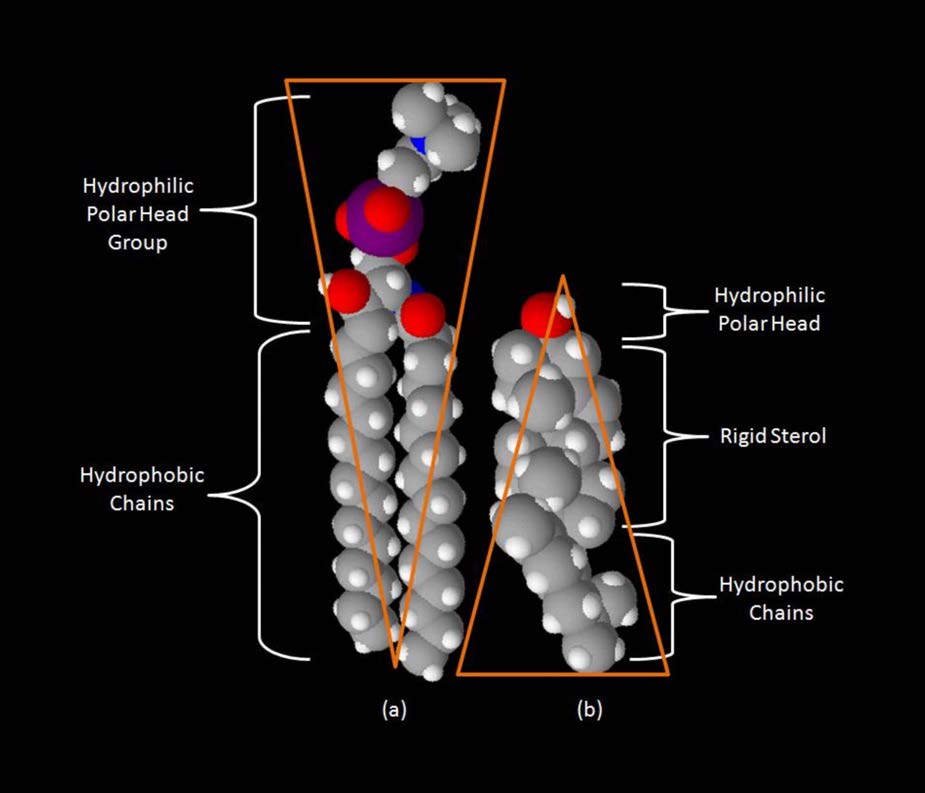
Aunque algunas células no tienen paredes celulares (células animales) y otras sí (bacterias, hongos y plantas), hay algo común entre las células en el sentido de que todas poseen membranas plasmáticas. También hay comunalidad en los componentes de las membranas, aunque la cantidad relativa de constituyentes varía. Las figuras 3.1 y 3.2 ilustran la estructura y los ambientes de las membranas plasmáticas. Todas las membranas plasmáticas contienen una cantidad significativa de sustancias anfifílicas ligadas a ácidos grasos. Estos incluyen los glicerofosfolípidos y los esfingolípidos. El (los) ácido (s) graso (s) se marcan como colas hidrófobas en las figuras.
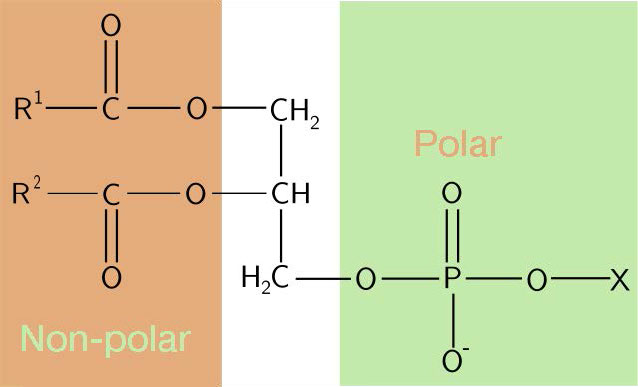
Cabezales hidrofílicos
La composición de las cabezas hidrofílicas varía considerablemente. En los glicerofosfolípidos, un fosfato siempre está presente, por supuesto, y a menudo se esterifica a otra sustancia para hacer un fosfátido (Figura 3.3). Los compuestos comunes unidos al fosfato (en la posición X) incluyen serina, etanolamina y colina. Estos varían en las sus cargas por lo que de esta manera, se puede controlar la carga en la superficie externa o interna. Las células tienden a tener más cargas negativas en la mitad exterior de la bicapa lipídica (llamada valva externa) y cargas más positivas en la mitad interior (folleto interno).
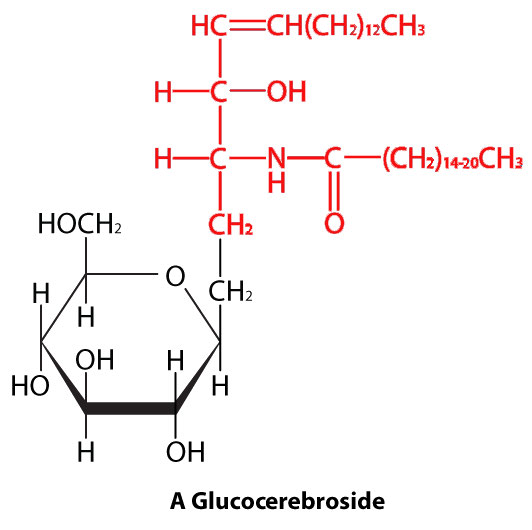
Esfingolípidos
En los esfingolípidos (Figura 3.4), la cabeza hidrófila puede contener un fosfato ligado a etanolamina o colina y esto describe la estructura de la esfingomielina, un componente importante de las membranas neurales. La mayoría de los esfingolípidos carecen del fosfato y en su lugar tienen una cabeza hidrofílica de un solo azúcar (cerebrosidos) o un oligosacárido complejo (gangliósidos).
Exclusión de agua
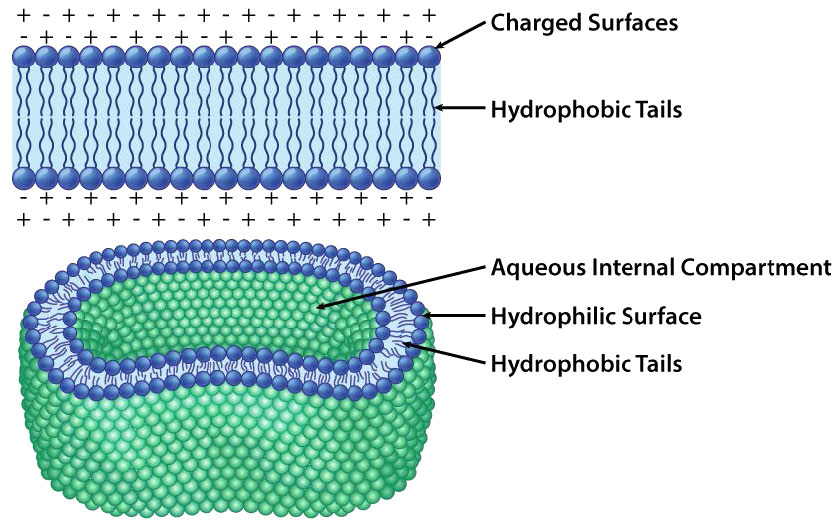
En cada caso, el glicerofosfolípido o esfingolípido tiene un extremo que es polar y un extremo que es no polar. Como vimos en la organización de aminoácidos con cadenas laterales hidrofóbicas que ocurren preferentemente en el interior de una proteína plegada para excluir el agua, también las porciones no polares de estas moléculas anfifílicas se disponen para excluir el agua. Recuerde que el citoplasma de una célula es principalmente agua y el exterior de la célula suele estar bañado en una capa acuosa. Por lo tanto, tiene perfecto sentido que las porciones polares de las moléculas de membrana se dispongan como lo hacen, partes polares en el exterior interactuando con el agua y partes no polares en el medio de la bicapa evitando o excluyendo el agua.
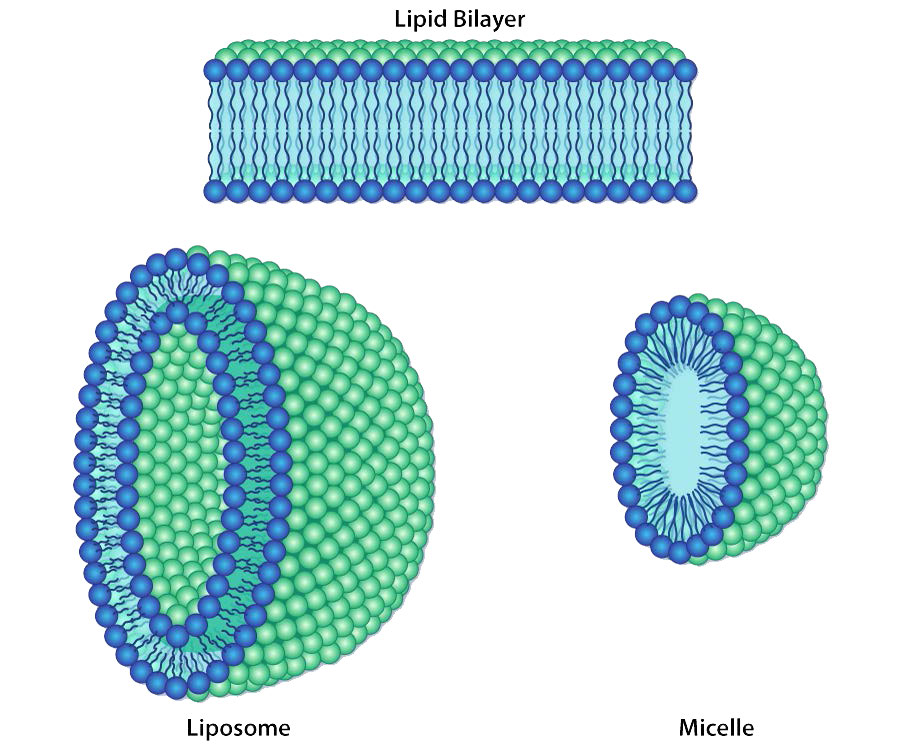
Sesgo de composición
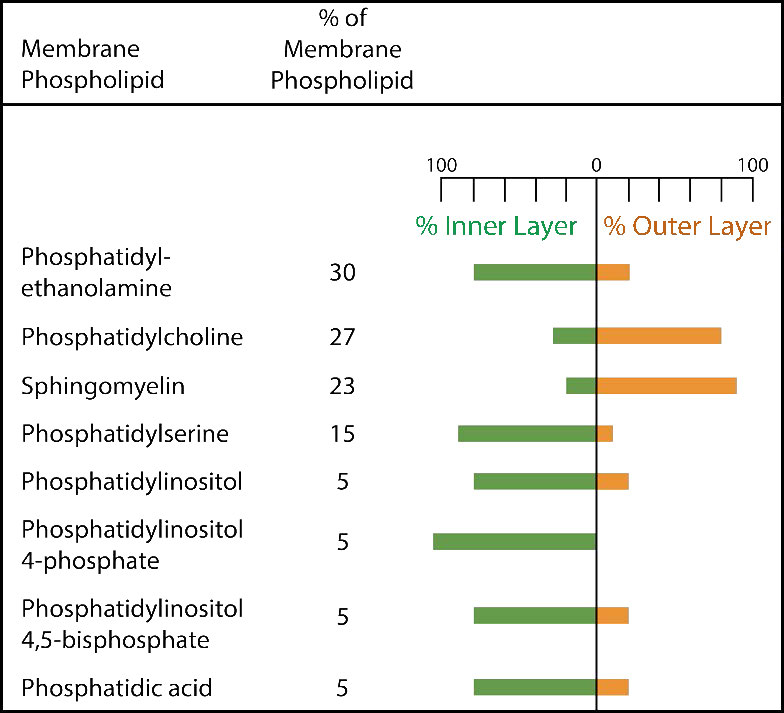
La membrana plasmática tiene distintos sesgos de composición en relación con su interior y exterior (Figura 3.7). En primer lugar, la glicosilación (de lípidos y proteínas) tiene los grupos de azúcar localizados casi exclusivamente en el exterior de la célula, alejados del citoplasma (Figura 3.8). Entre los lípidos de membrana, los esfingolípidos son mucho más comúnmente glicosilados que los glicerofosfolípidos. Además, algunos de los glierofosfolípidos se encuentran preferentemente en un lado u otro (Figura 3.7). La fosfatidilserina y la fosfatidiletanolamina se encuentran preferentemente dentro de la valva interna de la membrana plasmática, mientras que la fosfatidilcolina tiende a ubicarse en la valva externa. En el proceso de apoptosis, las fosfatidilserinas aparecen en la valva externa donde sirven como señal a los macrófagos para que se unan y destruyan la célula. Los esfingolípidos se encuentran preferentemente en la membrana plasmática y están casi completamente ausentes de las membranas mitocondriales y endoplásmicas del retículo (Figura 3.9).
Membranas organelares
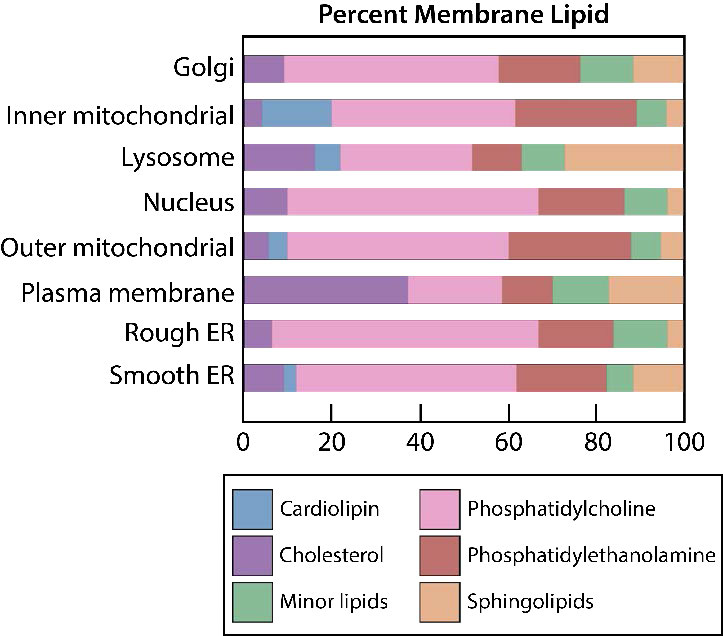
También existe sesgo de la composición lipídica con respecto a las membranas de orgánulos. El inusual difosfodiglicerolípido conocido como cardiolipina, por ejemplo, casi solo se encuentra en las membranas mitocondriales (ver AQUÍ) y al igual que la fosfatidilserina, su movimiento es un paso importante en la apoptosis. En la señalización, los fosfatidilinositoles desempeñan papeles importantes proporcionando segundos mensajeros al ser escindidos (ver AQUÍ).
Difusión Lateral
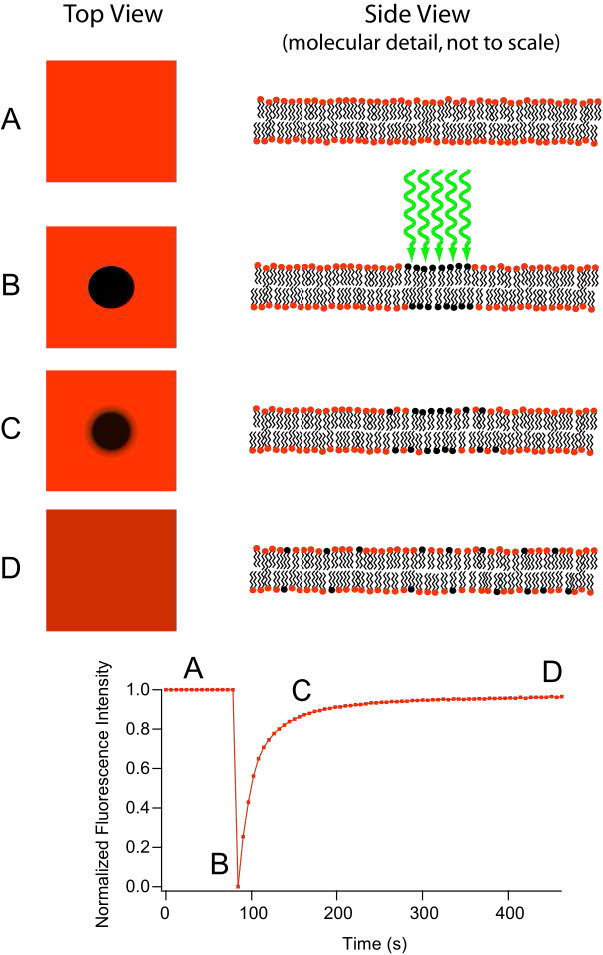
El movimiento de los lípidos dentro de cada valva de la bicapa lipídica ocurre fácil y rápidamente debido a la fluidez de la membrana. Este tipo de movimiento se denomina difusión lateral y se puede medir mediante la técnica denominada FRAP (Figura 3.10, ver AQUÍ también). En este método, un láser golpea y tiñe una sección de la bicapa lipídica de una célula, dejando una mancha como se muestra en B. Con el tiempo, la mancha se difunde finalmente a través de toda la bicapa lipídica, al igual que una gota de tinta se difundirá por todas partes cuando se agrega a un vaso de agua. Una medición de la velocidad de difusión da una indicación de la fluidez de una membrana.
Difusión Transversal
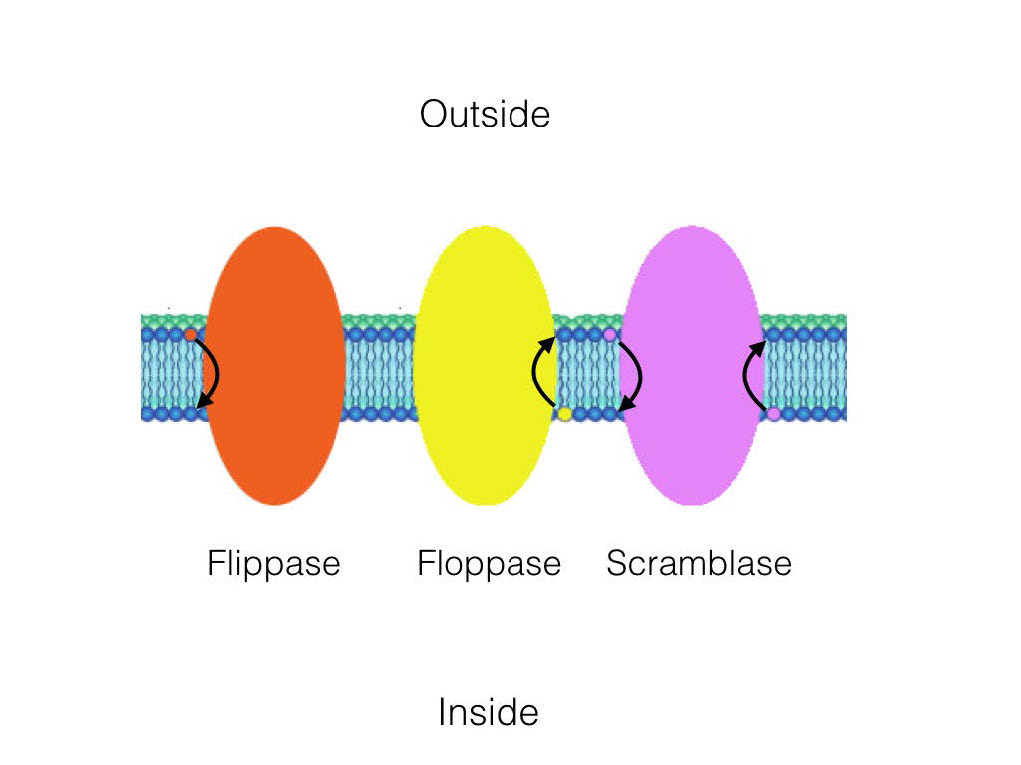
Mientras que el movimiento en la difusión lateral ocurre rápidamente, el movimiento de las moléculas de una valva a la otra se produce mucho más lentamente. Este tipo de movimiento molecular se denomina difusión transversal y es casi inexistente en ausencia de acción enzimática. Recuerde que existe un sesgo de distribución de moléculas entre las valvas de la membrana, lo que significa que algo debe estar moviéndolas.
Existen tres enzimas que catalizan el movimiento de compuestos en difusión transversal. Las flippasas mueven los glicerofosfolípidos/ esfingolípidos de membrana de la valva externa a la valva interna (lado citoplásmico) de la célula. Las floppasas mueven los lípidos de la membrana en dirección opuesta. Los Scramblases se mueven en cualquier dirección.
Otros componentes de la bicapa lipídica
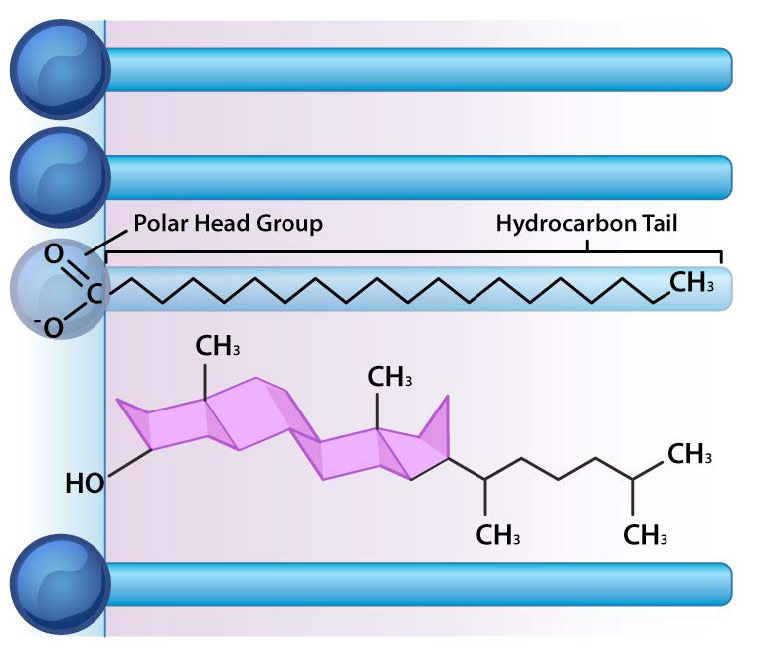
Además de glicerofosfolípidos y esfingolípidos, existen otros materiales comúnmente encontrados en bicapas lipídicas de membranas celulares. Dos importantes destacados son el colesterol (Figura 3.13) y las proteínas. Además de servir como precursor metabólico de las hormonas esteroides y los ácidos biliares, el papel principal del colesterol en las células está en las membranas. La planitud e hidrofobicidad de los anillos de esterol permiten que el colesterol interactúe con las porciones no polares de la bicapa lipídica mientras que el grupo hidroxilo en el extremo puede interactuar con la parte hidrófila.
Fluidez de la membrana
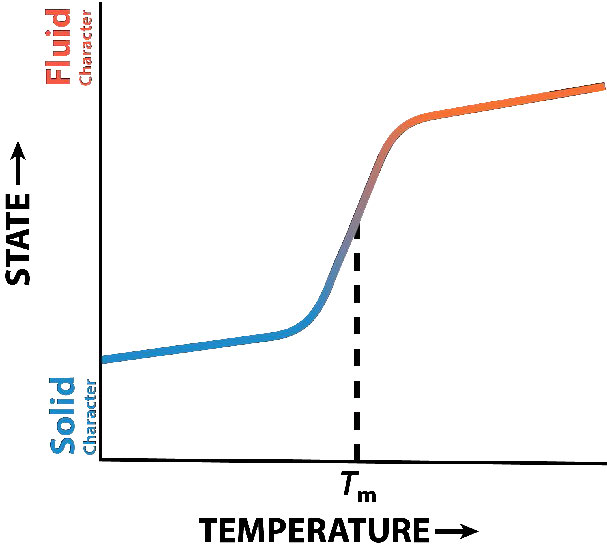
La función del colesterol en la bicapa lipídica es compleja (Figura 3.13). Influye en la fluidez de la membrana. La Figura 3.14 muestra la transición de fase para una membrana a medida que se calienta, pasando de un carácter más “congelado” al de uno más “fluido” a medida que aumenta la temperatura. El punto medio de esta transición, referido como la Tm, está influenciado por la composición de ácidos grasos de los compuestos de bicapa lipídica. Los ácidos grasos más largos y saturados favorecerán valores de Tm más altos, mientras que la insaturación y los ácidos grasos cortos favorecerán valores de Tm menores Es por esta razón que los peces, que viven en ambientes fríos, tienen un mayor nivel de ácidos grasos insaturados en ellos, para utilizarlos para elaborar lípidos de membrana que permanecerán fluidos a la temperatura del océano. Curiosamente, el colesterol no cambia el valor de Tm, sino que amplía el rango de transición entre las formas congeladas y fluidas de la membrana, lo que le permite tener un rango más amplio de fluidez.
Balsas lipídicas
El colesterol también se encuentra abundantemente en estructuras de membrana llamadas balsas lipídicas. Representadas en la Figura 3.15, las balsas lipídicas son estructuras organizadas dentro de la membrana que contienen típicamente moléculas de señalización y otras proteínas integrales de membrana. Las balsas lipídicas afectan la fluidez de la membrana, la neurotransmisión y el tráfico de receptores y proteínas de membrana.
Características
Las características distintivas de las balsas es que están más ordenadas que las bicapas que las rodean, contienen más ácidos grasos saturados (empaque más apretado y menos desorganización, como resultado) y hasta 5 veces más colesterol. También son relativamente ricos en esfingolípidos, con cantidades hasta 50% mayores de esfingomielina que las áreas circundantes de la bicapa. La mayor concentración de colesterol en las balsas puede deberse a su mayor capacidad de asociación con esfingolípidos (Figura 3.16). Algunos grupos, como las proteínas preniladas, como el RAS, pueden excluirse de las balsas lipídicas.
Las balsas lipídicas pueden proporcionar plataformas de concentración después de que los receptores de proteínas individuales se unen a ligandos en la señalización Después de que la activación del receptor tenga lugar en una balsa lipídica, el complejo de señalización proporcionaría protección contra las enzimas no balsas que podrían inactivar la señal. Por ejemplo, una característica común de los sistemas de señalización es la fosforilación, por lo que las balsas lipídicas podrían proporcionar protección contra la desfosforilación por enzimas llamadas fosfatasas. Las balsas lipídicas parecen estar involucradas en muchos procesos de transducción de señales, como la señalización del receptor de antígeno de células T, la señalización del receptor de antígeno de células B, la señalización del receptor de EGF, la señalización de inmunoglobulina E, la señalización del receptor Para más información sobre señalización, consulta AQUÍ.
Barrera
El transporte de materiales a través de membranas es esencial para que exista una célula. La bicapa lipídica es una barrera efectiva para la entrada de la mayoría de las moléculas y sin un medio para permitir que las moléculas de alimentos ingresen a una célula, moriría. Las moléculas primarias que se mueven libremente a través de la bicapa lipídica son pequeñas, sin carga, como H2O, CO2, CO y O2, por lo que las moléculas más grandes, como la glucosa, que la célula necesita de energía, serían efectivamente excluidas si no hubiera proteínas que facilitaran su movimiento a través de la membrana.
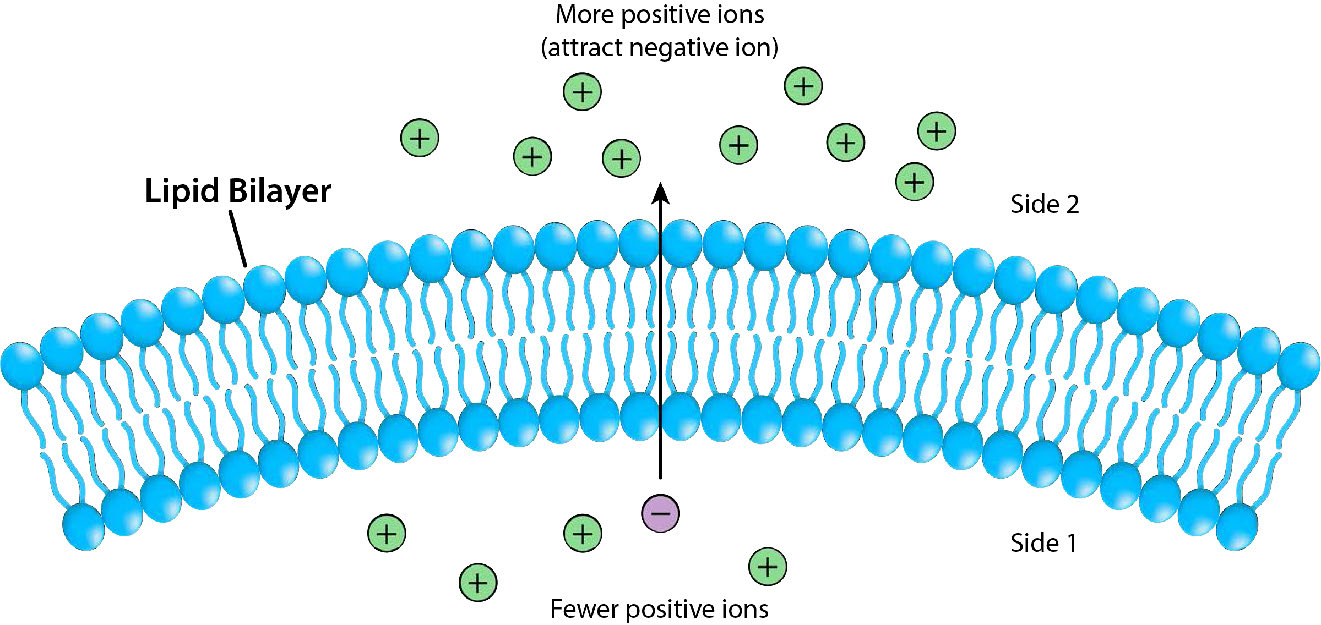
La Figura 3.17 representa la barrera que proporciona la bicapa lipídica al movimiento a través de ella y las presiones (atracción iónica, en este caso) que pueden afectar al movimiento. La energía potencial de las diferencias de carga y concentración es recolectada por las células para fines que incluyen la síntesis de ATP y el movimiento de materiales contra un gradiente de concentración en un proceso llamado transporte activo.
Proteínas de membrana
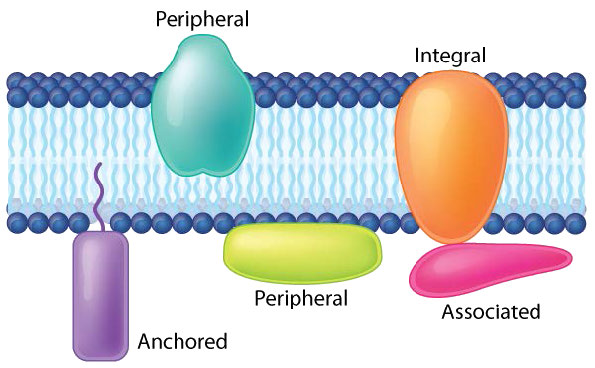
Las proteínas en una bicapa lipídica pueden variar enormemente en cantidad, dependiendo de la membrana. El contenido de proteínas en peso de varias membranas típicamente oscila entre 30 y 75% en peso. Algunas membranas mitocondriales pueden tener hasta 90% de proteína. Las proteínas ligadas y asociadas con las membranas vienen en varios tipos.
Proteínas transmembrana
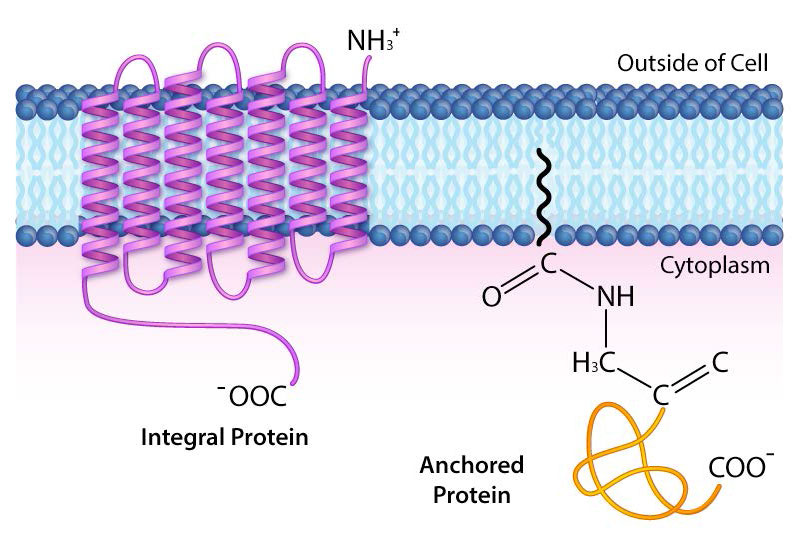
Las proteínas transmembrana son proteínas integrales de membrana que se extienden completamente de un lado de una membrana biológica al otro y están firmemente incrustadas en la membrana (Figura 3.18). Las proteínas transmembrana pueden funcionar como sitios de acoplamiento para la unión (a la matriz extracelular, por ejemplo), como receptores en el sistema de señalización celular, o facilitar el transporte específico de moléculas dentro o fuera de la célula.
Los ejemplos de proteínas integradas/transmembrana incluyen las implicadas en el transporte (por ejemplo, Na+/K+ ATPasa), canales iónicos (por ejemplo, canal de potasio de células nerviosas) y transducción de señales a través de la bicapa lipídica (por ejemplo, receptores acoplados a proteínas G).
Las proteínas de membrana periférica interactúan con parte de la bicapa (generalmente no implica interacciones hidrofóbicas), pero no se proyectan a través de ella. Un buen ejemplo es la fosfolipasa A2, que escinde los ácidos grasos de los glicerofosfolípidos en las membranas. Las proteínas de membrana asociadas normalmente no tienen regiones hidrofóbicas externas, por lo que no pueden incrustarse en una porción de la bicapa lipídica, sino que se encuentran cerca de ellas. Dicha asociación puede surgir como resultado de la interacción con otras proteínas o moléculas en la bicapa lipídica. Un buen ejemplo es la ribonucleasa.
Proteínas de membrana ancladas
Las proteínas de membrana ancladas no están ellas mismas incrustadas en la bicapa lipídica, sino que están unidas a una molécula (típicamente un ácido graso) que está incrustada en la membrana (Figura 3.19). La familia oncogénica de proteínas conocida como ras son buenos ejemplos. Estas proteínas se anclan a la bicapa lipídica mediante la unión a grupos farnesil no polares catalizados por la enzima farnesiltransferasa.
Clasificación más fina
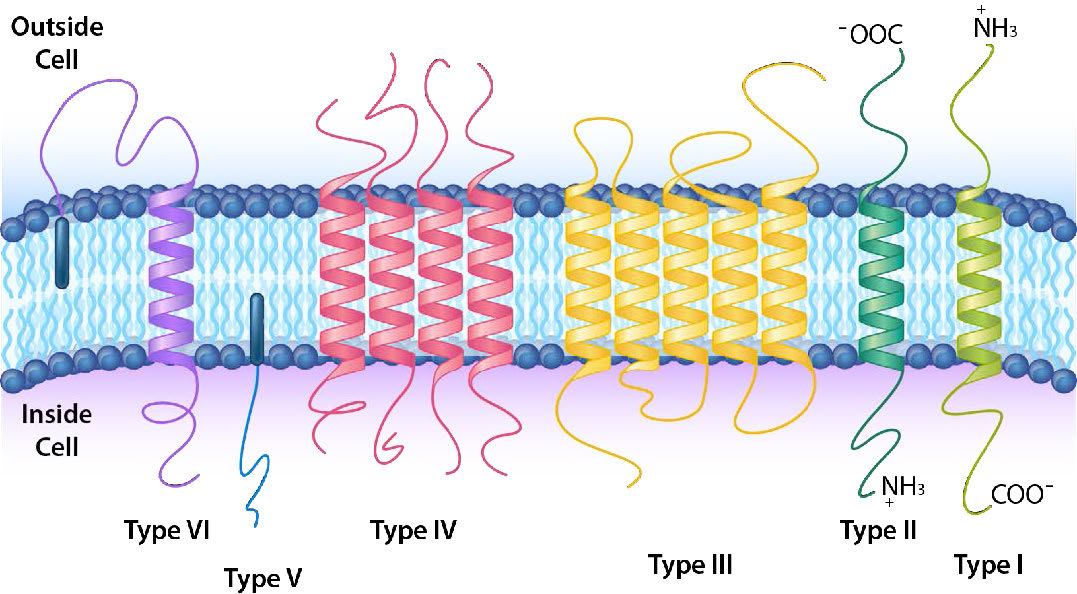
Un esquema de clasificación más detallado categoriza además las proteínas integrales y ancladas en seis tipos diferentes (Figura 3.20). El tipo I y el tipo II tienen solo una porción de la proteína que pasa a través de la membrana. Difieren en la orientación del extremo amina y carboxilo con respecto al interior/exterior. Las proteínas transmembrana tipo I tienen el extremo amino en el exterior y el extremo carboxi en el interior, mientras que las proteínas de tipo II tienen esto invertido. Las proteínas de tipo III son una sola cadena polipeptídica que tiene múltiples regiones que se cruzan hacia adelante y hacia atrás a través de la membrana, a menudo para formar un canal. El tipo IV es una proteína multipolipeptídica que tiene múltiples cruces de la membrana. Las proteínas transmembrana tipo V no tienen una parte de ellas que cruza la membrana, pero están ancladas a la membrana por un lípido (como un ácido graso) incrustado en la bicapa lipídica. Las proteínas transmembrana tipo VI tienen una porción de ellas que cruza la membrana y están unidas a un lípido incrustado en la bicapa lipídica.
Tipos de sangre
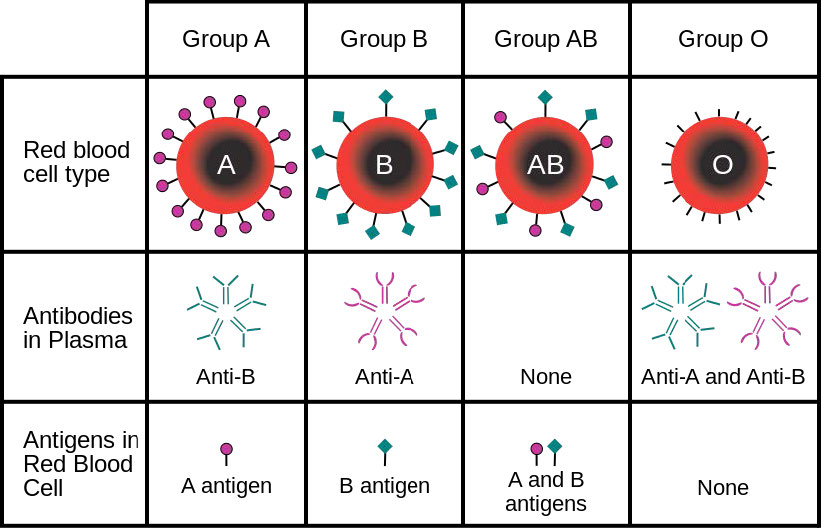
Las células tienen cientos de miles de proteínas de membrana y la composición proteica de una membrana varía según su función y ubicación. Las glicoproteínas incrustadas en las membranas juegan un papel importante en la identificación celular. Los tipos sanguíneos, por ejemplo, difieren entre sí en la estructura de las cadenas de carbohidratos que sobresalen de la superficie de la glicoproteína en sus membranas (Figura 3.21).
Presión osmótica
Las membranas proporcionan barreras/límites para la mayoría de las moléculas, pero la permeabilidad del agua a través de una bicapa lipídica crea una variable que debe considerarse. La variable aquí es la presión osmótica. La presión osmótica (floja) se refiere a la tendencia de una solución a absorber agua por el proceso de ósmosis. En la Figura 3.22, se puede observar una representación visual del concepto de la presión.
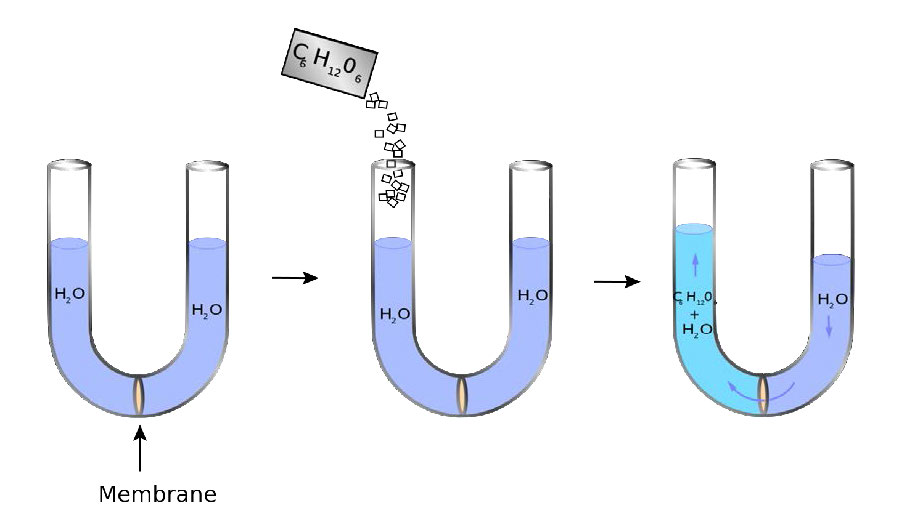
Un tubo en forma de U tiene en su parte inferior una membrana semipermeable. El agua puede pasar a través de la membrana, pero las moléculas de azúcar (C6H12O6) no pueden. En el lado izquierdo, se agrega azúcar creando una diferencia de concentración entre las cámaras derecha e izquierda. El agua se difunde a través de la membrana de derecha a izquierda en un intento de igualar las concentraciones, haciendo que el nivel del lado derecho disminuya y el lado izquierdo aumente. La presión resultante de las diferencias de altura se siente en la membrana.
Equivalación de concentraciones
Sin embargo, el líquido de la derecha no se mueve completamente hacia la izquierda, como podría esperarse si la única fuerza involucrada es igualar la concentración de azúcar a través de la membrana (sin azúcar a la derecha = sin agua). En cambio, se alcanza un equilibrio de tipos de niveles de agua a pesar de que las concentraciones no son iguales. La presión existente en la membrana luego de las diferencias de nivel corresponde a la presión osmótica de la mezcla. La presión osmótica de una solución es la diferencia de presión necesaria para detener el flujo de disolvente a través de una membrana semipermeable. La presión osmótica también se puede considerar como la presión requerida para contrarrestar la ósmosis. La pres osmótica- Figura 3.21 - Los tipos de sangre surgen de las glicoproteínas de la superficie celular Figura 3.22 - Presión osmótica. El agua se difunde hacia la izquierda para tratar de igualar la concentración de soluto. La presión realizada en la membrana en la figura correcta es la presión osmótica segura de que una solución diluida se comporta matemáticamente como la ley de gas ideal
\[P_{osmotic} = \dfrac{nRT}{V}\]
donde n es el número de moles, R es la constante del gas, T es la temperatura en Kelvin y V es el volumen.
Es más conveniente en soluciones para trabajar con molaridad, por lo que
\[P_{osmotic}= MR^* T\]
donde M es la molaridad de las moléculas disueltas, R* es la constante de gas expresada en (L atm)/(K mol), y T es la temperatura. La letra griega π se utiliza para referirse al término osmótico P, por lo que
\[Π = MR^* T\]
Recuerda al calcular la molaridad para incluir la molaridad de cada partícula. Por ejemplo, cuando se disuelve la sacarosa en solución, no se divide en partículas más pequeñas, por lo que
\[Molarity_{Particles} = Molarity_{Sucrose}\]
Sin embargo, para sales, como KOH, que forma dos iones en solución (K+ y OH-),
Partículas de Molaridad = 2* Molaridad KOH.
Consideración significativa
La presión osmótica es una consideración significativa para las células. Considera el hecho de que el agua puede moverse libremente a través de las membranas celulares, pero la mayor parte del contenido de la célula, como proteínas, ADN, iones, azúcares, etc., no puede. Segundo, la concentración de estos compuestos dentro de la célula es diferente a la concentración de ellos fuera de la célula. Tercero, dado que el agua puede moverse a través de la bicapa lipídica, tenderá a moverse en la dirección que tenderá a igualar el soluto.
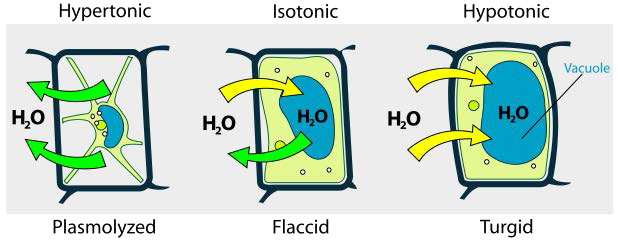
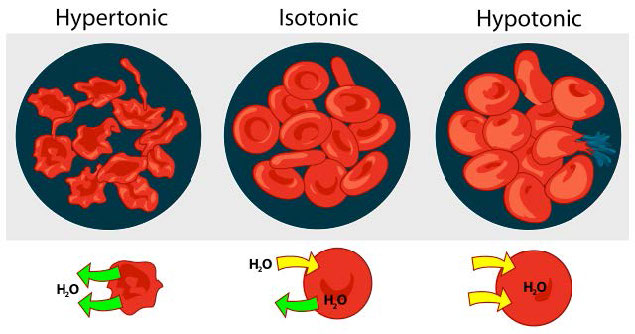
Hipotónico, hipertónico, isotónico
Consideramos tres situaciones (Figura 3.23). Primero, si la concentración de solutos es mayor dentro de la célula que en el exterior, el agua tenderá a moverse hacia la celda, haciendo que la célula se hinche. Esta circunstancia se llama hipotónica. Por el contrario, si la concentración de soluto es mayor fuera de la celda que dentro de ella, el agua saldrá de la celda y de la celda con encogimiento. Esta es una situación hipertónica. Por último, si las concentraciones de solutos dentro y fuera de la célula son iguales, a esto se le llama solución isotónica. Aquí, no se produce ningún movimiento de agua a través de la membrana celular y la célula conserva su tamaño.
Si la presión osmótica es mayor que las fuerzas que mantienen unida una membrana celular, la célula se romperá. Debido a esto, algunas células han incorporado defensas para evitar problemas. Las células vegetales, por ejemplo, tienen una pared celular bastante rígida que resiste la expansión en soluciones hipotónicas (Figura 3.24). Las bacterias también tienen una pared celular que brinda protección.