
5.1: Fundamentos de la Energía
- Page ID
- 53064

Fuente: BioChemFFA _5_1.pdf. Todo el libro de texto está disponible de forma gratuita de los autores en http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy

Los organismos vivos están formados por células, y las células contienen una horda de componentes bioquímicos. Las células vivas, sin embargo, no son colecciones aleatorias de estas moléculas. Están extraordinariamente organizados u “ordenados”. Por el contrario, en el mundo no vivo, existe una tendencia universal a aumentar el desorden. Mantener y crear orden en las celdas requiere la entrada de energía. Sin energía, la vida no es posible.
Energía Oxidativa
El principal mecanismo utilizado por los organismos no fotosintéticos para obtener energía es la oxidación y el carbono es la fuente de energía oxidada más comúnmente. La energía liberada durante las etapas oxidativas es “capturada” en ATP y puede ser utilizada posteriormente para el acoplamiento de energía. Cuanto más reducido es un átomo de carbono, más energía se puede obtener de su oxidación. Los ácidos grasos son muy reducidos, mientras que los carbohidratos lo son moderadamente. La oxidación completa de ambos conduce al dióxido de carbono, que tiene el estado energético más bajo. Por el contrario, cuanto más oxidado está un átomo de carbono, más energía se necesita para reducirlo.
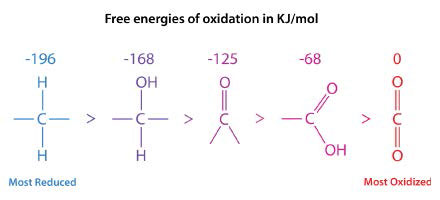
En la serie mostrada en la Figura\(\PageIndex{1}\), la forma más reducida de carbono se encuentra a la izquierda. La energía de oxidación de cada forma se muestra arriba de ella. Los estados de reducción de los ácidos grasos y carbohidratos se pueden ver por sus fórmulas.
- Ácido palmítico:\(\ce{C16H34O2}\)
- Glucosa:\(\ce{C6H12O6}\)
El ácido palmítico solo contiene dos oxígenos por dieciséis carbonos, mientras que la glucosa tiene seis átomos de oxígeno por seis carbonos. En consecuencia, cuando el ácido palmítico está completamente oxidado, genera más ATP por carbono (128/16) que glucosa (38/6). Es por esto que utilizamos la grasa (contiene ácidos grasos) como nuestro material de almacenamiento de energía primaria.
Figura\(\PageIndex{2}\): Fotosíntesis: La fuente primaria de energía biológica. Imagen de Aleia Kim
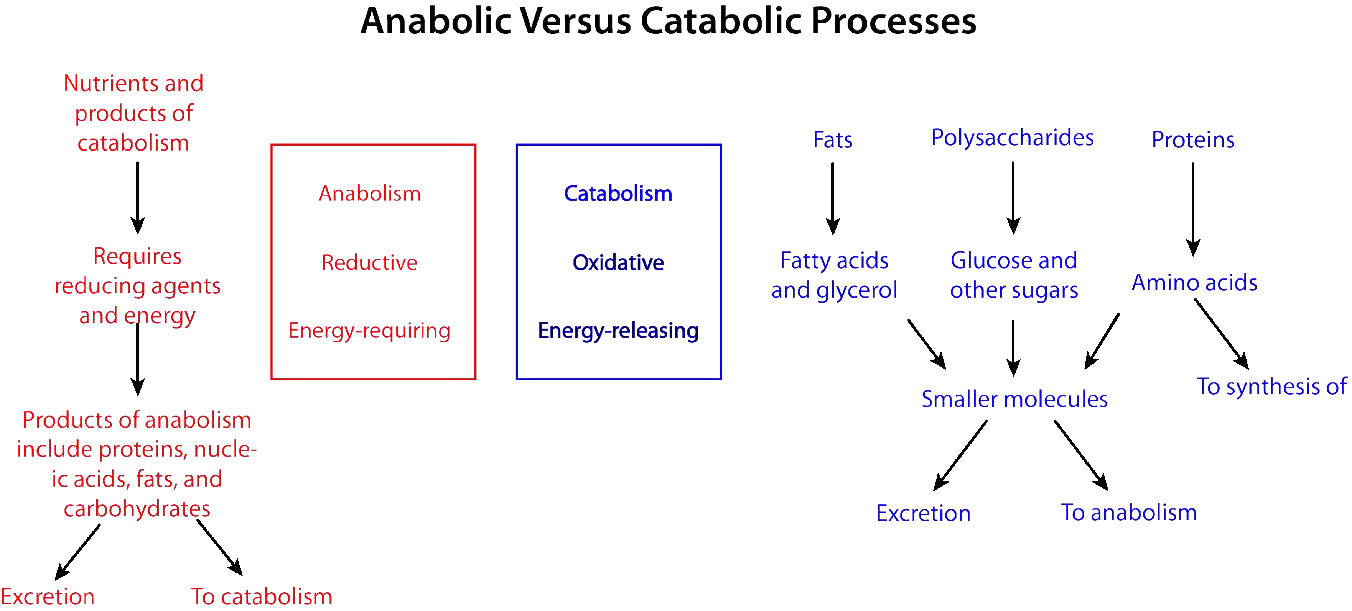
Oxidación vs Reducción del Metabolismo
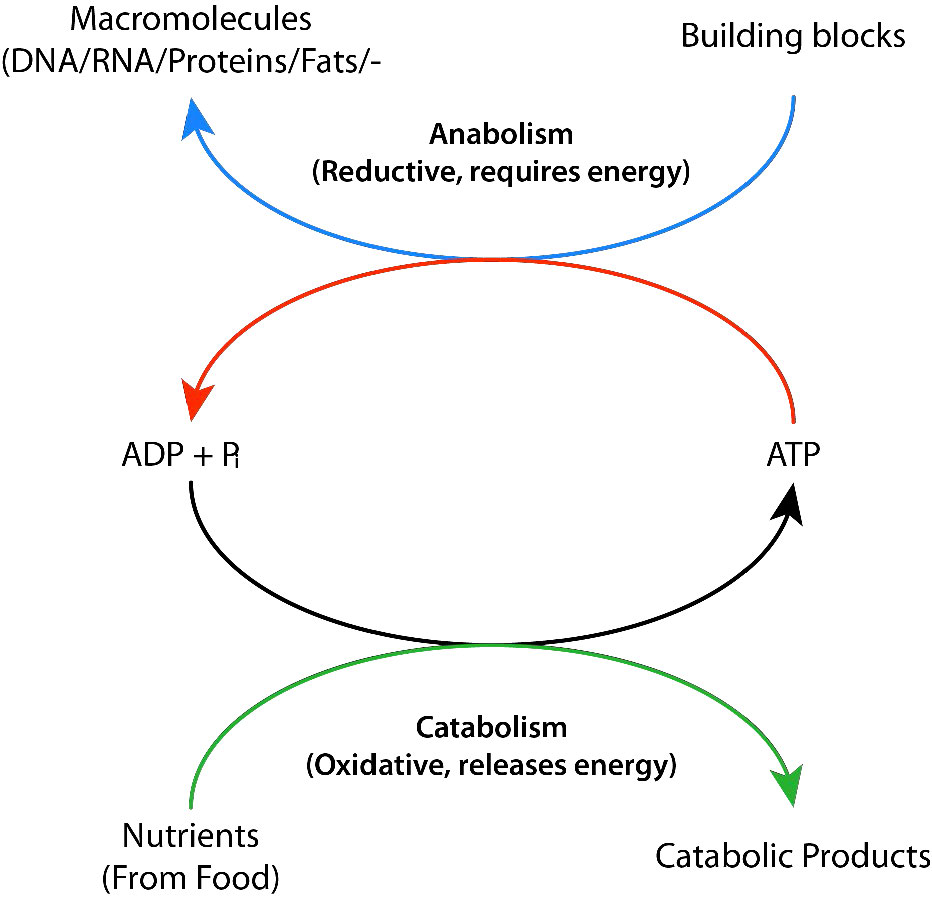
Los procesos bioquímicos que descomponen las cosas de más grandes a más pequeñas se denominan procesos catabólicos. Los procesos catabólicos suelen ser de naturaleza oxidativa y liberadores de energía. Parte, pero no toda, de esa energía es capturada como ATP. Si no toda la energía es capturada como ATP, ¿qué pasa con el resto? La respuesta es sencilla. Se libera como calor y es por ello que nos calentamos cuando hacemos ejercicio.
Por el contrario, sintetizar moléculas grandes a partir de otras más pequeñas (por ejemplo, hacer proteínas a partir de aminoácidos) se conoce como anabolismo. Los procesos anabólicos suelen ser de naturaleza reductora (Figuras 5.3 y 5.4) y requieren un aporte de energía. Por sí mismos, no se producirían, ya que están invirtiendo la oxidación y disminuyendo la entropía (convirtiendo muchas cosas pequeñas en una más grande). Para superar esta barrera energética, las células deben gastar energía. Por ejemplo, si se desea reducir\(\ce{CO2}\) a carbohidratos, se debe usar energía para hacerlo. Las plantas hacen esto durante las reacciones oscuras de la fotosíntesis (Figura\(\PageIndex{3}\)). La fuente de energía para la reducción es en última instancia el sol. Los electrones para la reducción provienen del agua, y el\(\ce{CO2}\) se retira de la atmósfera y se incorpora a un azúcar.
Acoplamiento de Energía
La síntesis de las muchas moléculas que necesitan las células necesita el aporte de energía para que ocurra. Las células superan este obstáculo energético mediante el uso de ATP para “impulsar” la reacción (Figura\(\PageIndex{6}\)). La energía necesaria para impulsar las reacciones se cosecha en condiciones muy controladas en enzimas. Esto implica un proceso llamado 'acoplamiento'. Las reacciones acopladas se basan en vincular una reacción energéticamente favorable (es decir, una con un ∆G°' negativo) con la reacción que requiere un aporte de energía, que tiene un ∆G°' positivo. Siempre que el ∆G°' global de las dos reacciones juntas sea negativo, la reacción puede continuar. La hidrólisis del ATP es una reacción muy favorable energéticamente que comúnmente se vincula a muchas reacciones que requieren energía en las células. Sin la hidrólisis de ATP (o GTP, en algunos casos), la reacción no sería factible.
Entropía y energía
La mayoría de los estudiantes que han tenido algo de química conocen la Segunda Ley de la Termodinámica con respecto al trastorno creciente de un sistema. Las células son estructuras muy organizadas u ordenadas, lo que lleva a algunos a concluir erróneamente que la vida de alguna manera viola la segunda ley. De hecho, esa noción es incorrecta. La segunda ley no dice que la entropía siempre aumenta, solo eso, dejada sola, tiende a hacerlo, en un sistema aislado. Las células no son sistemas aislados, sin embargo, en que obtienen energía, ya sea del sol, si son autótrofas, o alimentos, si son heterótrofas.
Para contrarrestar la tendencia universal hacia el desorden a escala local se requiere energía. Como ejemplo, tome una baraja de cartas fresca que esté perfectamente alineada con Ace-King-Queen.. 4,3,2 por cada palo. Lanza la baraja al aire, dejando que las cartas se dispersen. Cuando los recojas, estarán más desordenados que cuando empezaron. Sin embargo, si pasas unos minutos (y gastas un poco de energía), puedes reorganizar el mismo mazo de regreso a su estado anterior, organizado. Si la entropía siempre aumentaba en todas partes, no podrías hacer esto. No obstante, con el aporte de energía, superaste el desorden. Esto ilustra un concepto importante: el costo de combatir el desorden es la energía.
Energía biológica
Hay, por supuesto, otras razones por las que los organismos necesitan energía. Contracción muscular, síntesis de moléculas, neurotransmisión, señalización, termorregulación y movimientos subcelulares son ejemplos. ¿De dónde viene esta energía? Las monedas de energía son generalmente moléculas que contienen fosfato de alta energía. El ATP es el más conocido y más abundante, pero el GTP también es una fuente de energía importante (fuente de energía para la síntesis de proteínas). CTP participa en la síntesis de glicerofosfólidos y UTP se utiliza para la síntesis de glucógeno y otros compuestos de azúcar. En cada uno de estos casos, la energía está en forma de energía química potencial almacenada en los enlaces multifosfato. Hidrolizar esos enlaces libera la energía en ellos.
De los trifosfatos, el ATP es la fuente primaria de energía, actuando para facilitar la síntesis de los demás por acción de la enzima NDPK. El ATP se produce mediante tres tipos distintos de fosforilación: fosforilación oxidativa (en mitocondrias), fotofosforilación (en cloroplastos de plantas) y fosforilación a nivel de sustrato (en reacciones catalizadas enzimáticamente).
Gibbs energía libre en Biología
El ATP generalmente se considera la “batería de almacenamiento” de las células (Ver también 'Copias de seguridad de baterías moleculares para músculos AQUÍ). Para entender cómo se captura la energía, primero debemos entender la energía libre de Gibbs y al hacerlo, comenzamos a ver el papel de la energía en la determinación de las direcciones que toman las reacciones químicas.
La energía libre de Gibbs puede considerarse como la energía disponible para trabajar en un sistema termodinámico a temperatura y presión constantes. Matemáticamente, la energía libre de Gibbs se da como:
\[G = H – TS\]
donde\(H\) está la entalpía,\(T\) es la temperatura en Kelvin, y\(S\) es la entropía. A temperatura y presión estándar, cada sistema busca lograr un mínimo de energía libre. De esta manera, aumentar la entropía\(S\),, reducirá la energía libre de Gibbs. Del mismo modo, si se dispone de exceso de calor (reduciendo la entalpía\(H\)), también se puede reducir la energía libre.
Las células deben trabajar dentro de las leyes de la termodinámica, como se ha señalado, por lo que todas sus reacciones bioquímicas, también, están regidas por estas leyes. Ahora consideraremos la energía en la celda. El cambio en la energía libre de Gibbs (\(∆G\)) para una reacción es crucial, para ella, y por sí sola, determina si una reacción avanza o no.
\[∆G = ∆H – T ∆S.\]
Hay tres casos
- ∆G < 0: la reacción procede tal y como está escrito
- ∆G = 0: la reacción está en equilibrio
- ∆G > 0: la reacción transcurre a la inversa
Para una reacción
\[\ce{aA <=> bB}\]
(donde 'a' y 'b' son números enteros y A y B son moléculas) a pH 7, ∆G puede determinarse mediante la siguiente ecuación,
\[∆G = ∆G°’ + RT \ln(\frac{[B]^b}{[A]^a})\]
Para múltiples reacciones de sustrato, tales como
\[\ce{aA + cC <=> bB + dD}\]
\[∆G = ∆G°’ + RT \ln(\frac{[B]^b [D]^d}{[A]^a[C]^c})\]
El término ∆G°' se denomina cambio en la energía Standard Gibbs Free, que es el cambio en la energía que se produce cuando todos los productos y reactivos están en condiciones estándar y el pH es 7.0. Es una constante para una reacción dada.
En términos simples, podemos recopilar todos los términos del numerador juntos y llamarlos {Productos} y todos los términos del denominador juntos y llamarlos {Reactantes},
\[∆G = ∆G°’ + RT \ln(\frac{\rm{\{Products\}}}{\rm{\{Reactants\}}})\]
Para la mayoría de los sistemas biológicos, la temperatura, T, es una constante para una reacción dada. Dado que ∆G°' también es una constante para una reacción dada, el ∆G se cambia casi exclusivamente a medida que cambia la relación de {Productos}/{Reactantes}.
Importancia de ∆G°'
Si uno comienza en condiciones estándar, donde todo excepto los protones está en 1M, el término rTLn ({Productos}/{Reactivos}) es cero, por lo que el término ∆G°' es igual al ∆G, y el ∆G°' determina la dirección que tomará la reacción (solo bajo esas condiciones). Es por ello que la gente dice que un ∆G°' negativo indica una reacción energéticamente favorable, mientras que un ∆G°' positivo corresponde a una desfavorable.
Al aumentar la relación de {Productos}/{Reactivos}, el valor del término logaritmo natural (ln) se vuelve más positivo (menos negativo), haciendo así que el valor de ∆G sea más positivo. Por el contrario, a medida que disminuye la relación de {Productos}/{Reactivos}, el valor del término logarítmico natural se vuelve menos positivo (más negativo), haciendo así que el valor de ∆G sea más negativo.
Respuesta del sistema al estrés
Intuitivamente, esto tiene sentido y es consistente con el Principio de Le Chatelier: un sistema responde al estrés actuando para aliviar el estrés. Si examinamos el ∆G para una reacción en un sistema cerrado, vemos que siempre se moverá a un valor de cero (equilibrio), no importa si comienza con un valor positivo o negativo.
Otro tipo de energía libre disponible para las células es la generada por el potencial eléctrico. Por ejemplo, las mitocondrias y los cloroplastos utilizan en parte energía coulómbica (basada en la carga) de un gradiente de protones a través de sus membranas para proporcionar la energía necesaria para la síntesis de ATP. Energías similares impulsan la transmisión de señales nerviosas (gradientes de sodio y potasio) y el movimiento de algunas moléculas en procesos de transporte activo secundario a través de las membranas (por ejemplo, el diferencial H+ que impulsa el movimiento de la lactosa). De la ecuación de cambio de energía libre de Gibbs,
\[∆G = ∆H – T∆S\]
cabe señalar que un aumento en la entropía ayudará a contribuir a una disminución de ∆G Esto ocurre, por ejemplo cuando una molécula grande se está rompiendo en trozos más pequeños o cuando el reordenamiento de una molécula aumenta el trastorno de las moléculas a su alrededor. Esta última situación surge en el efecto hidrofóbico, que ayuda a impulsar el plegamiento de las proteínas.
Potencial químico y eléctrico
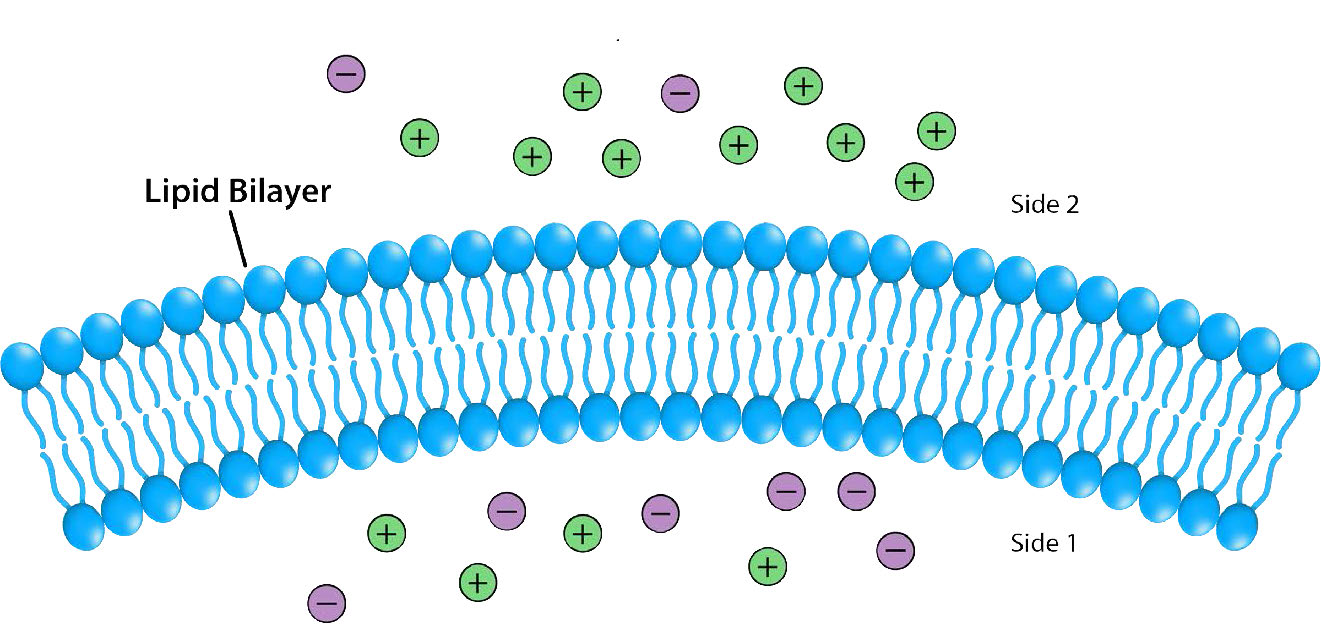
Se dice que la ausencia hace crecer el cariño al corazón. Aquí no abordaremos ese tema filosófico, pero diremos que la separación proporciona energía potencial que las células pueden cosechar y hacen. La bicapa lipídica de las membranas celulares y (en células eucariotas) de orgánulos proporcionan la barrera necesaria para la separación.
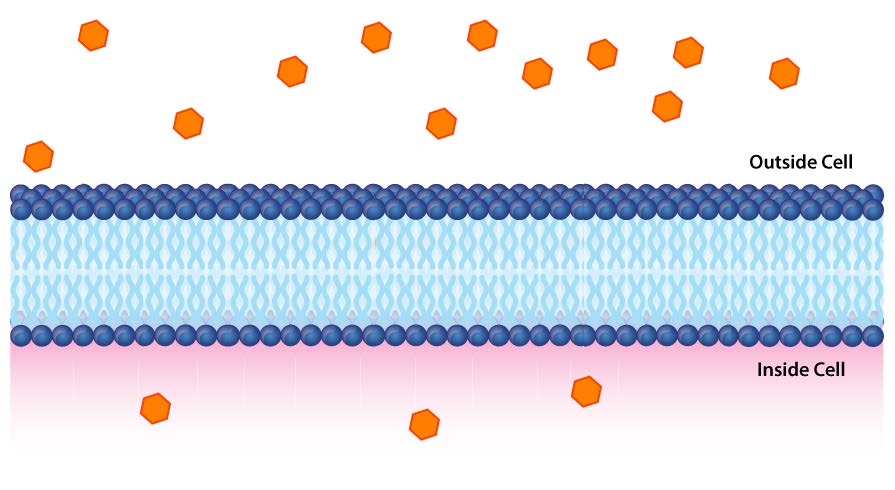
Impermeables a la mayoría de los iones y compuestos polares, las membranas biológicas son esenciales para los procesos que generan energía celular. Considere la Figura 5.8. Una bicapa lipídica separa dos soluciones con diferentes concentraciones de un soluto. Hay una mayor concentración de iones negativos en la parte inferior y una mayor concentración de iones positivos en la parte superior.
Siempre que hay una diferencia en la concentración de moléculas a través de una membrana, se dice que hay un gradiente de concentración a través de ella. Una diferencia en la concentración de iones a través de una membrana también crea un gradiente de carga (o eléctrico). Debido a que existe una diferencia tanto en la concentración química de los iones como en la carga en los dos lados de la membrana, esto se describe como un gradiente electroquímico (Figuras 5.8 -5.10).
Energía potencial
Dichos gradientes funcionan como baterías y contienen energía potencial. Cuando las células cosechan la energía potencial, pueden crear ATP, transmitir señales nerviosas, bombear moléculas a través de membranas y más. Es importante, por lo tanto, entender cómo calcular la energía potencial de los gradientes electroquímicos.
Primero, consideramos gradientes químicos (solutos). En la Figura 5.9, dos concentraciones de glucosa están separadas por una bicapa lipídica. Supongamos que C2 es la concentración de glucosa dentro de la célula (abajo) y C1 la concentración de glucosa fuera (arriba). La energía libre de Gibbs asociada con el movimiento de la glucosa en la dirección de C2 (hacia la célula) viene dada por
∆G = RTLn [C2/C1]
Para moverlo en la dirección de C1 (hacia el exterior de la celda) la expresión sería
\[∆G = RT\ln[C_1/C_2]\]
Dado que C2 es menor que C1 (es decir, hay menos moléculas de glucosa dentro de la célula) entonces el ∆G es negativo y la difusión se favorecería en la célula, si la glucosa pudiera atravesar la bicapa.
Por el contrario, si C2 fuera mayor que C1 (había más glucosa en la célula que en el exterior) el ∆G sería positivo, por lo que no se favorecería el movimiento en la dirección de C2 y en su lugar la glucosa tendería a moverse hacia C1, es decir, fuera de la célula.
Si C2 = C1, con la misma concentración de glucosa dentro y fuera, entonces el ∆G sería cero y no habría movimiento neto, ya que el sistema estaría en equilibrio.
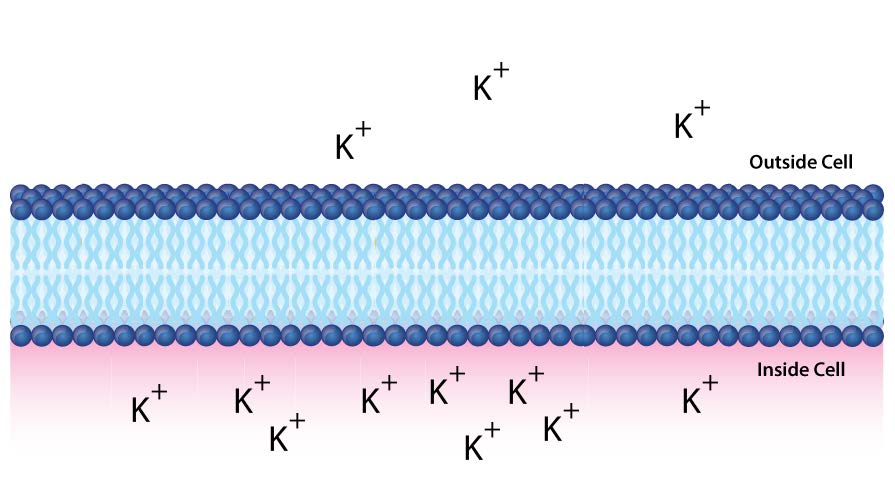
En el ejemplo anterior, consideramos la glucosa, que es una molécula sin carga. Cuando están involucrados iones, también se deben tomar en consideración sus cargas. La Figura\(\PageIndex{1}\) 0 representa una situación similar a través de una bicapa lipídica. En este caso, existe una diferencia de concentración y carga. Hay más cargas positivas dentro de la celda que afuera.
Usando C2 para indicar la concentración de materiales dentro de la celda y C1 para la concentración fuera de la celda (como antes), entonces la energía libre para el movimiento de un ion de arriba a abajo viene dada por la siguiente ecuación
\[∆G = RT\ln[C_2/C_1] + ZF∆ψ\]
Obsérvese aquí que esta ecuación debe tomar en consideración tanto las diferencias de concentración como las diferencias de carga. Z se refiere a la carga de las especies transportadas, F es la constante de Faraday (96,485 culombs/mol) y ∆ψ es la diferencia de potencial eléctrico (diferencia de voltaje) a través de la membrana.
Si calculáramos el ∆G para el movimiento del ion potasio de arriba a abajo, sería positivo, ya que [C2/C1] es mayor que 1 (haciendo para un término ln positivo), y el Zf∆ψ es positivo porque los iones cargados positivamente (Z) se mueven contra un gradiente de carga positiva dado por ∆ψ (mayor concentración en el objetivo (abajo) que en el punto de partida (arriba)). Si calculáramos la concentración de iones moviéndose de abajo hacia arriba, entonces el término ln sería negativo (C2<C1) y el Zf∆ψ también sería negativo (Z=positivo, pero ∆ψ negativo).
Potencial de reducción
Al discutir el potencial químico, también debemos considerar el potencial de reducción. El potencial de reducción mide la tendencia de una sustancia química a ser reducida por electrones. También es designado por varios otros nombres/variables. Estos incluyen potencial redox, potencial de oxidación/reducción, ORP, pE, ε, E y Eh.
El potencial de reducción se mide en voltios o milivoltios. Una sustancia con un mayor potencial de reducción tendrá una mayor tendencia a aceptar electrones y ser reducida. Si se mezclan dos sustancias en una solución acuosa, la que tenga el mayor potencial de reducción (más positivo) tenderá a quitar electrones, reduciéndose así, de la que tiene el menor potencial de reducción, que se oxida.
Medidas relativas
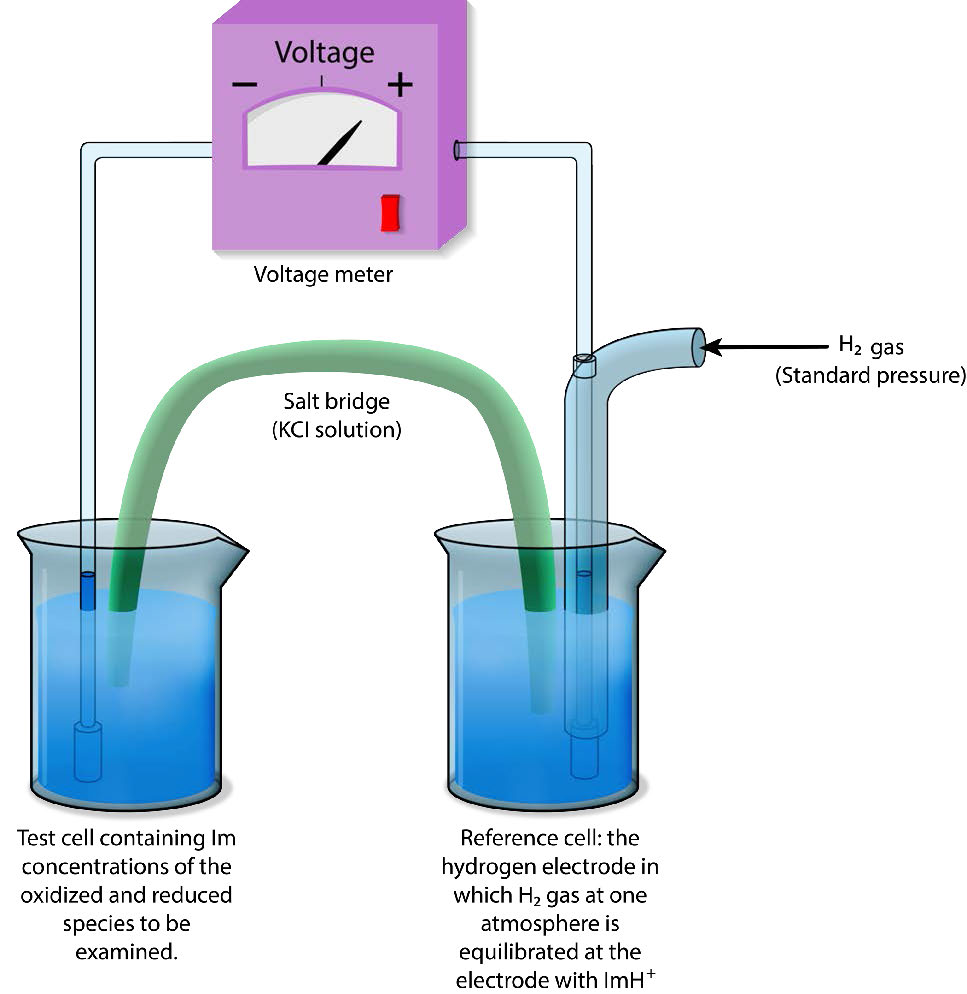
Los potenciales de reducción absolutos son difíciles de medir, por lo que los potenciales de reducción se definen típicamente en relación con un electrodo de referencia. En soluciones acuosas, los potenciales de reducción se miden como la diferencia de potencial entre un electrodo sensor inerte (típicamente platino) en contacto con la solución de prueba y un electrodo de referencia estable (medido como un Electrodo de Hidrógeno Estándar: SHE) como se muestra en la Figura\(\PageIndex{1}\) 1. El estándar de referencia para la medición es la media reacción
H + e — → ½ H 2
Al electrodo donde se produce esta reacción (denominado semicelda) se le da el valor de E° (Potencial de Reducción Estándar) de 0.00 voltios. El electrodo de hidrógeno se conecta a través de un circuito externo a otra media celda que contiene una mezcla de las especies reducidas y oxidadas de otra molécula (por ejemplo, Fe ++ y Fe +++) a 1M cada una y condiciones estándar de temperatura (25°C) y presión (1 atmósfera).
Dirección y voltaje medidos
Luego se mide la dirección y magnitud del movimiento de electrones. Si la mezcla de prueba toma electrones del electrodo de hidrógeno, el signo del voltaje es positivo y si se invierte la dirección, el voltaje es negativo.
Así, los compuestos que tienen mayor afinidad por los electrones que el hidrógeno registrarán un voltaje positivo y los voltajes negativos corresponden a compuestos con menor afinidad por los electrones que el hidrógeno.
Movimiento de electrones
Bajo condiciones estándar, los electrones pasarán de los compuestos que generan voltajes más bajos a los que generan voltajes más altos (más positivos). Así como el cambio de energía libre de Gibbs estándar es el cambio de energía libre de Gibbs en condiciones estándar, también lo es el potencial de reducción estándar E° el potencial de reducción E en condiciones estándar.
El potencial de reducción real de una media célula variará con la concentración de cada especie química en la célula. La relación entre el potencial de reducción E y el potencial de reducción estándar E° viene dada por la siguiente ecuación (también llamada ecuación de Nernst)
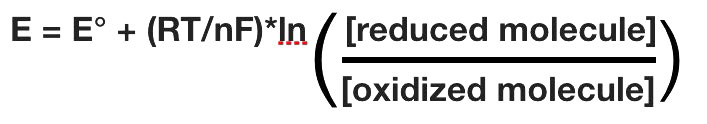
donde F es la constante de Faraday (96,480 J/ (voltios*moles), R es la constante de gas (8.315 J/ (moles*K), n es el número de moles de electrones que se transfieren, y T es la temperatura absoluta en Kelvin.
A 25°C, esta ecuación se convierte en
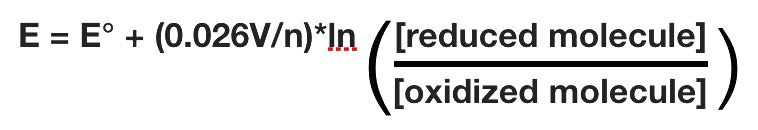
En cuanto a la energía libre de Gibbs, es útil medir valores en condiciones que se encuentran en las células. Esto significa hacer mediciones a pH = 7, lo que difiere de tener todas las especies a 1M.
Ajuste
Debido a este ajuste, se define un potencial de reducción estándar ligeramente diferente y lo designamos por E°', así como definimos un cambio de energía libre de Gibbs estándar especial a pH 7 como ΔG°'.
Existe una relación entre el cambio en la energía libre de Gibbs ΔG y el cambio en el potencial de reducción (ΔE). Es
\[ΔG = -nFΔE\]
De igual manera, la relación entre el cambio en la energía libre estándar de Gibbs y el cambio en el potencial de reducción estándar es
\] ΔG°' = -NFδE°'\]
Almacenamiento de Energía en Trifosfatos
Película 5.1: ATP: El combustible de la celda
La formación de trifosfatos, como el ATP, es esencial para satisfacer las necesidades energéticas de la célula para la síntesis, el movimiento y la señalización. En un día dado, un cuerpo humano promedio produce y descompone más que su peso en trifosfatos. Esto es especialmente notable considerando que solo hay alrededor de 250 g de la molécula presente en el cuerpo en un momento dado. La energía en ATP es liberada por hidrólisis de un fosfato de la molécula.
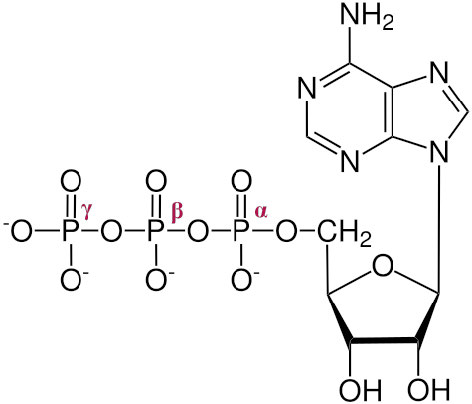
Los tres fosfatos, comenzando por el más cercano al azúcar, se denominan α, β y γ (Figura\(\PageIndex{1}\) 2). Es el γ fosfato el que se escinde en la hidrólisis y el producto es ADP. En algunas reacciones, se escinde el enlace entre α y β. Cuando esto sucede, se libera un pirofosfato (β ligado a γ) y se produce AMP. Esta última reacción para producir AMP libera más energía (ΔG°' = -45.6 kJ/mol) que la primera reacción que produce ADP (ΔG°' = -30.5 kJ/mol).
Dado que los trifosfatos son la “moneda” que satisface las necesidades inmediatas de la célula, es importante entender cómo se hacen los trifosfatos. Existen tres mecanismos de fosforilación: 1) nivel de sustrato; 2) oxidativo; y 3) fotofosforilación. Los consideramos aquí de manera individual.
Fosforilación a nivel de sustrato
El tipo de fosforilación más fácil de entender es el que ocurre a nivel de sustrato. Este tipo de fosforilación implica la síntesis directa de ATP a partir de ADP y un intermedio de alta energía, típicamente una molécula que contiene fosfato. La fosforilación a nivel de sustrato es un contribuyente relativamente menor a la síntesis total de trifosfatos por las células. Un ejemplo de fosforilación del sustrato proviene de la glucólisis.
Fosfoenolpiruvato (PEP) + ADP Piruvato + ATP
Esta reacción tiene un ∆G°' muy negativo (-31.4 kJ/mol), lo que indica que el PEP contiene más energía que ATP, tendiendo así a favorecer energéticamente la síntesis de ATP. También se pueden hacer otros trifosfatos por fosforilación a nivel de sustrato. Por ejemplo, el GTP se puede sintetizar mediante la siguiente reacción del ciclo del ácido cítrico.
succinil-CoA + PIB + Pi succinato + GTP + CoA-SH
Los trifosfatos pueden intercambiarse fácilmente en fosforilaciones a nivel de sustrato catalizadas por la enzima Nucleósido Difosfato quinasa (NDPK). Una forma generalizada de las reacciones catalizadas por esta enzima es la siguiente:
XTP + YDP XDP + YTP
donde X = adenosina, citidina, uridina, timidina o guanosina e Y puede ser cualquiera de estas también. Además, XTP e YDP también pueden ser cualquiera de los desoxinucleotidos.
Por último, una forma inusual de sintetizar ATP por fosforilación a nivel de sustrato es a través de la reacción catalizada por la adenilato quinasa
2 ADP ATP + AMP
Fuente ATP
Esta reacción es un medio importante para generar ATP cuando la célula no tiene otras fuentes de energía. La acumulación de AMP resultante de esta reacción activa enzimas, como la fosfofructoquinasa, de la glucólisis, que catalizarán reacciones para dar a la célula la energía adicional necesaria.
Es importante señalar que las enzimas no pueden hacer que ocurran reacciones que sean energéticamente desfavorables. Las enzimas aceleran las reacciones, pero no cambian su dirección. Las células están así ligadas por las reglas de la energía libre de Gibbs. Entonces, ¿cómo ocurren reacciones energéticamente desfavorables en una célula?
Acoplamiento de reacción
Las reacciones que son energéticamente desfavorables, se pueden hacer favorables acoplándolas con la hidrólisis de ATP, una reacción muy energéticamente favorable. Existen numerosos paralelismos en el “mundo real”. El movimiento de los automóviles es energéticamente desfavorable, pero el acoplamiento del movimiento del automóvil a la oxidación de la gasolina hace que un proceso desfavorable sea favorable. Otro enfoque para hacer favorable una reacción desfavorable es manipular la concentración de reactivos y productos. Considere la reacción a continuación, que ocurre en el metabolismo de nucleótidos de pirimidina:
orotato + PRPP OMP + PP i
El ΔG°' para esta reacción es -0.8 kJ/mol, lo que significa que si se inicia con concentraciones iguales de reactivos y productos, en equilibrio, habrá un pequeño exceso de productos. En la celda, sin embargo, esta reacción se mueve fuertemente hacia la derecha (ΔG = muy negativo). Dado que el ΔG°' es muy cercano a cero, solo puede ocurrir un ΔG muy negativo si se alteran las concentraciones de reactivos y productos, ya que
\[ΔG = ΔG°’ + RT \ln(\frac{[\rm{OMP}][\rm{PP_i}]}{[\rm{Orotate}][\rm{PRPP}]})\]
La manipulación es exactamente lo que sucede aquí. El ítem clave cuya concentración se ajusta en esta reacción es el pirofosfato (PPi). Esto es posible porque las células contienen una enzima llamada pirofosforilasa que cataliza la siguiente reacción
PP i + H 2 O 2 P i
La hidrólisis del pirofosfato es muy favorecida energéticamente, haciendo que el PPi producido en la reacción se hidroliza rápidamente. Como resultado, la concentración de PPi en la célula se mantiene muy baja. Una baja concentración de un producto (PPi) hace que el término logarítmico natural (ln) de la ecuación orotada se vuelva más negativo, lo que impulsa el término ΔG para que la reacción general se vuelva mucho más negativa.
Empujar y tirar
Las reacciones que producen pirofosfato como producto se producen en la síntesis de ADN y ARN, así como muchas otras moléculas. Como se muestra en el ejemplo anterior, este pirofosfato se hidroliza rápidamente, haciendo que la reacción general se mueva en la dirección de la producción de pirofosfato. Cuando los reactivos se eliminan/reducen en una reacción metabólica para disminuir la concentración de un producto, decimos que la reacción se “tira”, para representar el aumento en la reacción directa como resultado del agotamiento del producto.
El empuje ocurre cuando se añaden/aumentan los reactivos en una reacción. Esto también tiene el efecto de reducir el ΔG de una reacción y hacerla más favorable porque la relación de [Productos]/[Reactivos] disminuye con el aumento de [Reactivos]. Empujar y tirar de las reacciones son herramientas adicionales para que las células superen las barreras energéticas, al igual que acoplar procesos energéticamente favorables a otros energéticamente desfavorables.