7.4: Reparación de ADN
- Page ID
- 53000

Fuente: BiochemFFA_7_3.pdf. Todo el libro de texto está disponible gratuitamente de los autores en http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy

Salvaguardar el genoma
En la última sección se consideraron las formas en que las células afrontan los desafíos asociados a replicar su ADN, un proceso vital para todas las células. Es evidente que si el ADN es la copia maestra de instrucciones para un organismo, entonces es importante no cometer errores al copiar el ADN para pasarlo a nuevas células. Aunque la corrección de pruebas por ADN polimerasas aumenta en gran medida la precisión de la replicación, existen mecanismos adicionales en las células para garantizar aún más que el ADN recién replicado sea una copia fiel del original, y también para reparar el daño al ADN durante la vida normal de una célula.
Daño en el ADN
Todo el ADN sufre daños a lo largo del tiempo, por exposición a radiación ultravioleta y otras radiaciones, así como por diversos químicos en el ambiente (Figuras 7.34 y 7.35). Incluso las reacciones químicas que ocurren naturalmente dentro de las células pueden dar lugar a compuestos que pueden dañar el ADN. Como ya sabrás, incluso cambios menores en la secuencia del ADN, como las mutaciones puntuales, a veces pueden tener consecuencias de largo alcance. Asimismo, los daños no reparados causados por la radiación, los químicos ambientales o incluso la química celular normal pueden interferir con la transmisión precisa de la información en el ADN. Mantener la integridad del “plano” celular es de vital importancia y esto se refleja en los numerosos mecanismos que existen para reparar errores y daños en el ADN.
Reparación de desajuste post-replicativo
Anteriormente discutimos la corrección de pruebas por ADN polimerasas durante la replicación. Si bien la corrección reduce significativamente la tasa de error, no todos los errores son fijados sobre la marcha por las ADN polimerasas.
¿Qué mecanismos existen para corregir los errores de replicación que se pierden por la función correctora de ADN polimerasas? Los errores que se deslizan al corregir durante la replicación se pueden corregir mediante un mecanismo llamado reparación de desajustes. Si bien la tasa de error de replicación del ADN es de aproximadamente uno de cada 107 nucleótidos en ausencia de reparación de desapareamientos, esto se reduce aún más cien veces a uno de cada 109 nucleótidos cuando la reparación de desapareamientos es funcional.
¿Cuáles son las tareas a las que se enfrenta un sistema de reparación de desajuste
Debe:
- Escanee ADN recién hecho para ver si hay bases mal ventiladas (por ejemplo, una G emparejada con una T)
- Identificar y eliminar la región del desajuste.
- Llenar correctamente el vacío creado por la escisión de la región de desajuste.
Distinción de hebras
Es importante destacar que el sistema de reparación de desapareamientos debe tener un medio para distinguir la cadena de ADN recién hecha de la cadena molde, si los errores de replicación se van a corregir correctamente. En otras palabras, cuando el sistema de reparación de desajuste encuentra un mal juego A-G, por ejemplo, debe saber si la A debe ser removida y reemplazada por una C o si la G debe ser removida y reemplazada por una T.
Pero, ¿cómo distingue el sistema de reparación de desajustes entre las hebras originales y las nuevas de ADN? En bacterias, la existencia de un sistema que metila el ADN en las secuencias GATC es la solución a este problema. E. coli tiene una enzima, ADN adenina metilasa (Dam) que agrega grupos metilo en el a adeninas en secuencias GATC en ADN (Figura 7.36). El ADN recién replicado aún no ha sufrido metilación y, por lo tanto, se puede distinguir de la cadena molde, que está metilada.
Las proteínas reparadoras de desapareamientos reemplazan selectivamente la cadena que carece de metilación, asegurando así que son errores en la cadena recién hecha los que se eliminan y reemplazan. Debido a que la metilación es el criterio que permite que el sistema de reparación de desapareamientos elija la cadena que se repara, el sistema de reparación de desapareamientos bacterianos se describe como dirigido
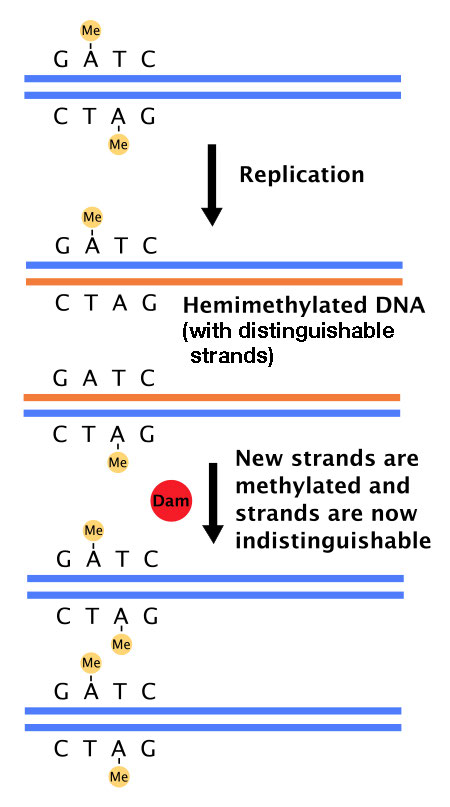
Figura 7.36- La metilasa Dam añade grupos metilo en las secuencias GATC
Genes de reparación de des
La reparación de desapareamientos ha sido bien estudiada en bacterias y se han identificado las proteínas involucradas. En E. coli, las proteínas reparadoras de desapareamientos son codificadas por un grupo de genes conocidos colectivamente como genes mut. Componentes importantes de la maquinaria de reparación de desapareamientos son las proteínas MuTs, MuTl y MuTh (Figura 7.37).
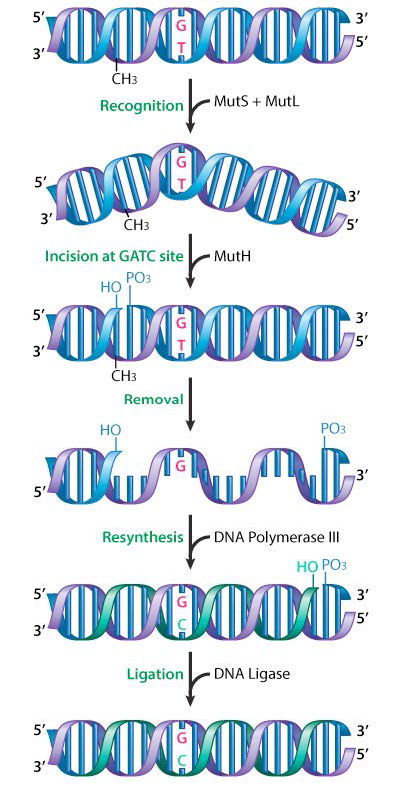
MuTs actúa para reconocer el desapareamiento, mientras que MuTl y MuTH son reclutados en el sitio de desapareamiento por la unión de MuT. MuTH es una endonucleasa que corta la cadena de ADN recién sintetizada y, hasta ahora, no metilada en un GATC. Esto activa una helicasa de ADN y una exonucleasa que ayudan a desenrollar y eliminar la región que contiene el desapareamiento. La ADN polimerasa III rellena el hueco, usando la hebra opuesta como molde, y la ligasa se une a los extremos, para restaurar una cadena continua.
Los eucariotas también tienen un sistema de reparación de desajustes que repara no solo los desajustes de base única sino también las inserciones y eliminaciones. Se han identificado homólogos de E. coli MuTs y mutL en otros organismos, incluyendo humanos: hMSH1 y hMSH2 (homólogo humano MuTs 1 y 2) son homólogos a MuTs, mientras que hMLH 1 es homólogo a MuTl. Estas, junto con proteínas adicionales, llevan a cabo la reparación de desapareamientos en células eucariotas.
La metilación del ADN no es utilizada por las células eucariotas como una forma de distinguir la nueva cadena del molde, y aún no se entiende completamente cómo el sistema de reparación de desapareamientos en eucariotas “sabe” qué cadena reparar. Existe evidencia de que el ADN recién hecho puede ser reconocido por el hecho de que es mellado, o discontinuo. Esto sugiere que la discontinuidad resultante de los fragmentos de Okazaki que aún no se han unido puede permitir que la nueva hebra se distinga de la antigua hebra plantilla continua.
Reparación de daños en el ADN
En la sección anterior observamos los errores cometidos cuando se copia el ADN, donde se inserta la base equivocada durante la síntesis de la nueva hebra. Pero incluso el ADN que no se está replicando puede dañarse o mutarse. Este tipo de daños no están asociados con la replicación del ADN, sino que pueden ocurrir en cualquier momento.
¿Qué causa daño al ADN?
Algunas de las principales causas de daño al ADN son:
a. Radiación (por ejemplo, rayos UV en la luz solar y en cabinas de bronceado, o radiación ionizante)
b. Exposición a sustancias químicas dañinas, como nitrosaminas o hidrocarburos aromáticos policíclicos, en el ambiente (ver Figura 7.38)
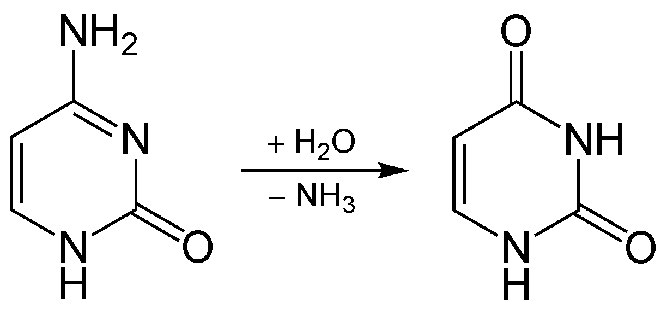
c. Reacciones químicas dentro de la célula (como la desaminación de citosina para dar uracilo, o la metilación de guanina para producir metilguanina).
Esto significa que el ADN en tus células es vulnerable al daño simplemente por tipos normales de acciones, como caminar al aire libre, estar en el tráfico o por las transformaciones químicas que ocurren en cada célula como parte de sus actividades cotidianas. (Naturalmente, el daño es mucho peor en situaciones donde la exposición a la radiación o productos químicos dañinos es mayor, como cuando las personas usan camas de bronceado, o fuman, regularmente).
Tipos de daños
¿Qué tipo de daño causan estos agentes? La radiación puede causar diferentes tipos de daño al ADN.
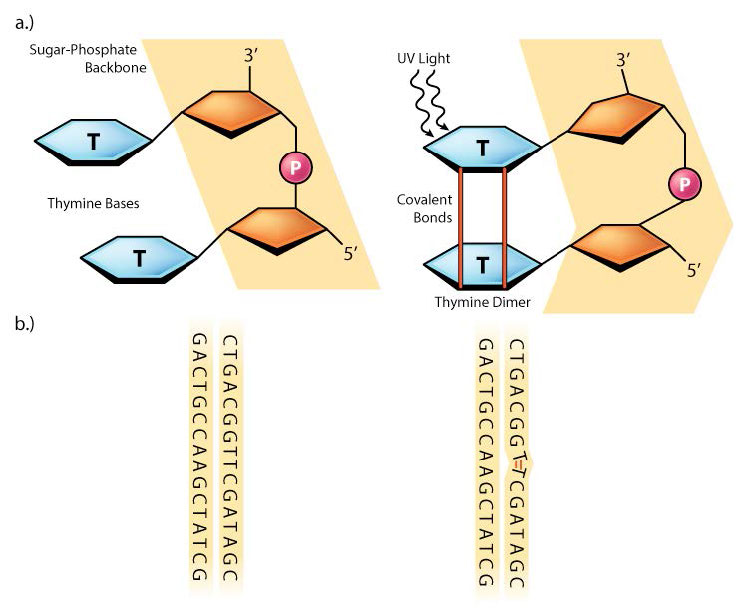
En ocasiones, como con gran parte del daño causado por los rayos UV, dos bases de pirimidina adyacentes en el ADN se reticularán para formar dímeros de ciclobutano pirimidina o CPD (ver Figura 7.39). Tenga en cuenta que estas son dos bases de pirimidina vecinas en la misma cadena de ADN. La exposición UV también puede conducir a la formación de otro tipo de lesión, conocida como fotoproducto (6-4) o 6-4PP (Figura 7.39). La radiación ionizante puede provocar roturas en la cadena principal del ADN, en una o ambas hebras.
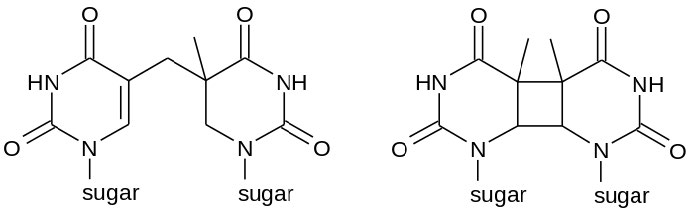
Figura 7.39 - Posibles estructuras químicas de un dímero de pirimidina - 6-4PP (izquierda) y CPD (derecha) - Wikipedia
Moléculas como el benzopireno, que se encuentran en los gases de escape de los automóviles, pueden unirse a bases, formando aductos de ADN voluminosos en los que grandes grupos químicos están unidos a bases en el ADN. Los daños como dímeros de pirimidina, 6-4PP o aductos químicos pueden distorsionar físicamente la hélice del ADN, provocando que la ADN y la ARN polimerasa se bloqueen cuando intentan copiar esas regiones del ADN (Figura 7.40).
Las reacciones químicas que ocurren dentro de las células pueden hacer que las citosinas en el ADN se desaminen a uracilo. Otros tipos de daño en esta categoría incluyen la formación de bases oxidadas como 8-oxo-guanina o bases alquiladas como O6-metilguanina. Estos en realidad no cambian la estructura física de la hélice del ADN, pero pueden causar problemas porque el uracilo y la 8-oxo-guanina se emparejan con bases diferentes a la citosina o guanina originales, lo que lleva a mutaciones en la siguiente ronda de replicación. De manera similar, la O6-metilguanina puede formar pares de bases con timina en lugar de citosina.
Eliminación de daños
Las celdas tienen varias formas de eliminar los tipos de daños descritos anteriormente. El primero de ellos se describe como inversión directa. Muchos organismos (aunque, desafortunadamente para nosotros, no los humanos) pueden reparar el daño UV como los CPD y 6-4PP porque poseen enzimas llamadas fotóliasas (foto=luz; liasa = enzima de descomposición - Figura 7.41). Las fotóliasas trabajan a través de un proceso llamado fotoreactivación, y utilizan energía de luz azul para catalizar una reacción fotoquímica que rompe los enlaces aberrantes en el ADN dañado y devuelve el ADN a su estado original.
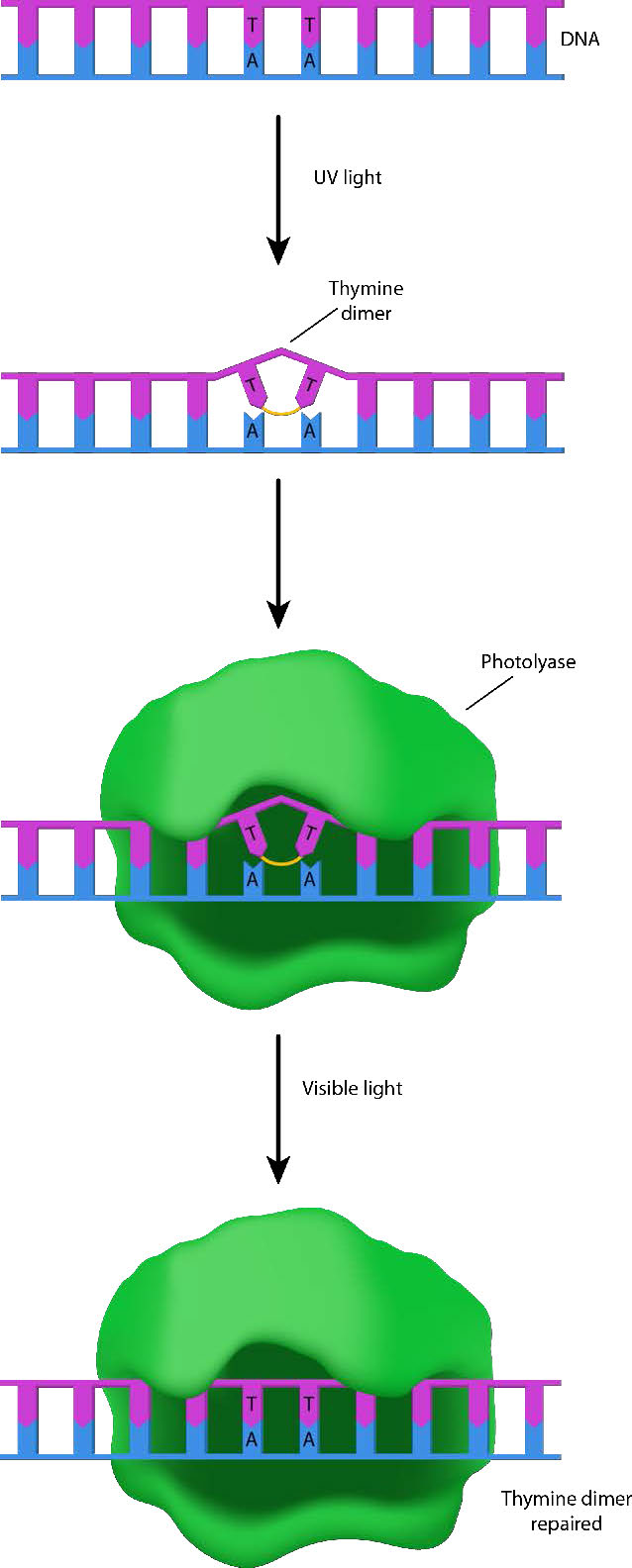
Enzima suicida
La O6-metilguanina en el ADN también se puede eliminar por reversión directa, con la ayuda de la enzima O6-metilguanina metiltransferasa. Esta es una enzima muy inusual que elimina el grupo metilo de la guanina y lo transfiere a un residuo de cisteína en la enzima. La adición del grupo metilo a la cisteína hace que la enzima no sea funcional.
Como saben, la mayoría de las enzimas son catalizadores que permanecen sin cambios durante el transcurso de la reacción, permitiendo que una sola molécula enzimática catalice repetidamente una reacción. Debido a que la O6-metilguanina metiltransferasa no se ajusta a esta descripción, a veces no es considerada como una verdadera enzima. También se le ha llamado enzima suicida, porque la enzima “muere” como resultado de su propia actividad.
Reparación de escisión
La reparación por escisión es otra estrategia común. Reparación por escisión es un término general para cortar y resintetizar la región dañada de un ADN. Existen varios tipos diferentes de reparación por escisión, pero todos implican extirpar la porción del ADN que está dañada, seguido de síntesis de reparación usando la otra cadena como molde, y finalmente, ligadura para restaurar la continuidad a la cadena reparada. Las células poseen varios tipos diferentes de reparación por escisión, cada una orientada a tipos específicos de daño en el ADN. Entre ellos, estos sistemas de reparación se ocupan de la amplia variedad de insultos al genoma.
Reparación de escisión de nucleótidos
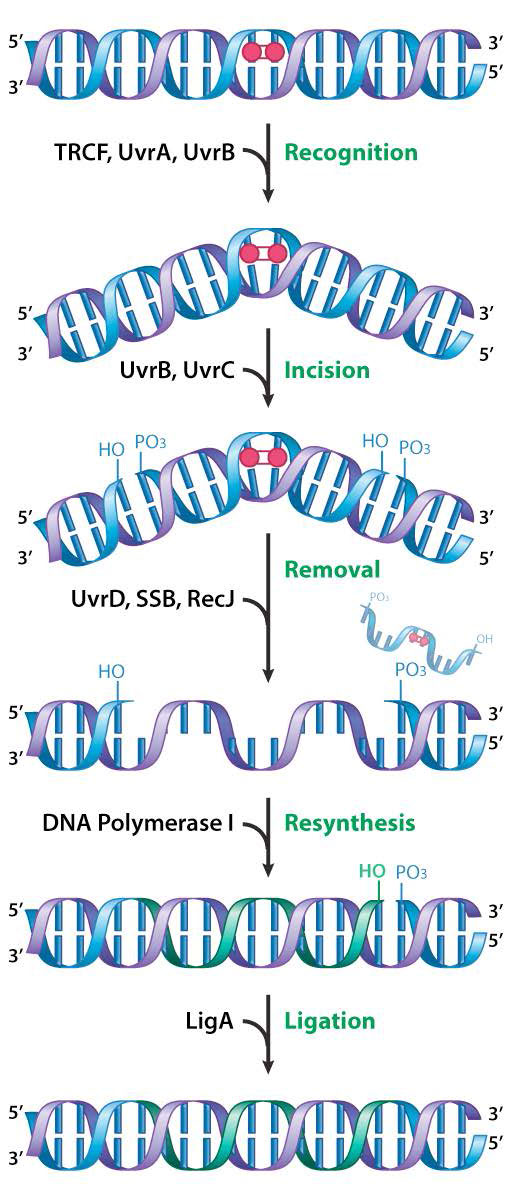
La reparación por escisión de nucleótidos (NER) corrige daños como la formación de aductos químicos, así como daños UV. Tanto los aductos químicos como la formación de CPD o 6,4 fotoproductos pueden causar distorsión significativa de la hélice del ADN. Las proteínas NER actúan para cortar la hebra dañada a ambos lados de la lesión. Luego se elimina una porción corta de la cadena de ADN que contiene el daño y una ADN polimerasa rellena el hueco con los nucleótidos apropiados. La reparación por escisión de nucleótidos ha sido ampliamente estudiada en bacterias.
En E. coli, el reconocimiento y escisión del daño se lleva a cabo por un grupo de proteínas codificadas por los genes uvrabC y uvRD. Los productos proteicos de los genes uvrA, uvrB y uvrC funcionan juntos como la llamada excinucleasa uvrabC. El daño es inicialmente reconocido y unido por un complejo de las proteínas UvrA y UvrB. Una vez unido el complejo, el UvrA se disocia, dejando el UvrB unido al ADN, donde luego se une por la proteína UvrC.
mellado de hilo
Es el complejo que consiste en UvrB y C el que actúa para cortar el esqueleto fosfodiéster a cada lado del daño, creando mellas en la cadena con una separación de aproximadamente 12-13 nucleótidos. Una helicasa codificada por UvrD luego desenrolla la región que contiene el daño, desplazándola de la doble hélice junto con UvrBC. La brecha en el ADN se rellena con la ADN polimerasa, que copia la hebra no dañada, y la mella se sella con la ayuda de la ADN ligasa.
La reparación por escisión de nucleótidos también es una vía importante en eucariotas. Es particularmente importante en la eliminación del daño UV en humanos, dado que carecemos de fotóliasas. Se han identificado varias proteínas que funcionan de manera similar a las proteínas Uvr.
La importancia de estas proteínas es evidente por el hecho de que las mutaciones en los genes que las codifican pueden conducir a una serie de enfermedades genéticas, como Xeroderma pigmentoso, o XP. Las personas con XP son extremadamente sensibles a la exposición a los rayos UV, debido a que el daño que provoca no se puede reparar, dejándolas en un riesgo mucho mayor de desarrollar cáncer de piel.
Dos modos de reparación
La reparación por escisión de nucleótidos opera en dos modos, uno conocido como reparación genómica global y el otro como reparación acoplada a transcripción. Si bien la función de ambos es eliminar daños desestabilizadores de hélice como dímeros de ciclobutano pirimidina o aductos químicos, la forma en que se detectan las lesiones difiere.
En la reparación genómica global, el daño se identifica mediante la vigilancia de todo el genoma para detectar lesiones que distorsionan la hélice. En el caso de la reparación acoplada a la transcripción, el bloqueo de la ARN polimerasa en un sitio de daño en el ADN es el indicador que activa este modo de reparación por escisión de nucleótidos.
Reparación de escisión de base
La reparación por escisión de base (BER) es un mecanismo de reparación que trata situaciones como la desaminación de citosina a uracilo (Figura 7.43) o la metilación de una base purina. Estos cambios no suelen distorsionar la estructura de la hélice del ADN, a diferencia de los aductos químicos o el daño UV.
En la reparación por escisión de bases, primero se elimina una sola base dañada del ADN, seguido de la eliminación de una región del ADN que rodea la base faltante. Luego se repara la brecha.
Uracil-ADN glicosilasa
La eliminación del uracilo del ADN se logra mediante la enzima uracil-ADN glicosilasa que puede reconocer uracilo en el ADN y romper el enlace glicosídico entre el uracilo y el azúcar en el nucleótido (Figura 7.44). La remoción de la base deja un hueco llamado sitio apirimidínico (sitio AP) porque, en este caso, se eliminó uracilo, una pirimidina. Es importante recordar que en este punto la columna vertebral del ADN sigue intacta, y la eliminación de una sola base simplemente crea un hueco como un diente que ha sido noqueado.
La formación del sitio AP desencadena la actividad de una enzima conocida como endonucleasa AP que corta el esqueleto del ADN 5' al sitio AP. En los pasos restantes, una ADN polimerasa se une a la mella, luego usando sus actividades de exonucleasa y polimerasa, reemplaza la secuencia en esta región. Dependiendo de la situación, se puede reemplazar un solo nucleótido (BER de parche corto) o puede eliminarse y reemplazarse un tramo de varios nucleótidos (BER de parche largo). Finalmente, como siempre, la ADN ligasa actúa para sellar la mella en el ADN.
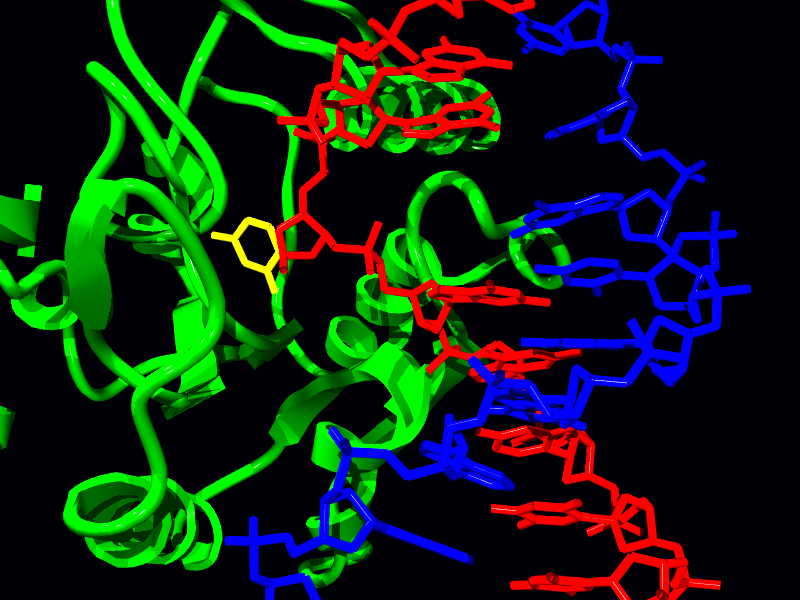
Reparación de roturas de doble hebra
Si bien todos los mecanismos de reparación discutidos hasta ahora fijaron el daño en una cadena de ADN usando la otra cadena no dañada como molde, estos mecanismos no pueden reparar el daño a ambas cadenas. ¿Qué pasa si ambos hilos están dañados? La radiación ionizante, la exposición a ciertos productos químicos o las especies reactivas de oxígeno generadas en la célula pueden provocar roturas bicatenarias (DSB) en el ADN.
Las DSB son una forma potencialmente letal de daño que, además de bloquear la replicación y transcripción, también puede conducir a translocaciones cromosómicas, donde parte de un cromosoma se adhiere a una pieza de otro cromosoma. Existen dos mecanismos celulares diferentes que ayudan a reparar las DSB (Figura 7.45), la recombinación homóloga (HR) y la unión de extremos no homólogos (NHEJ).
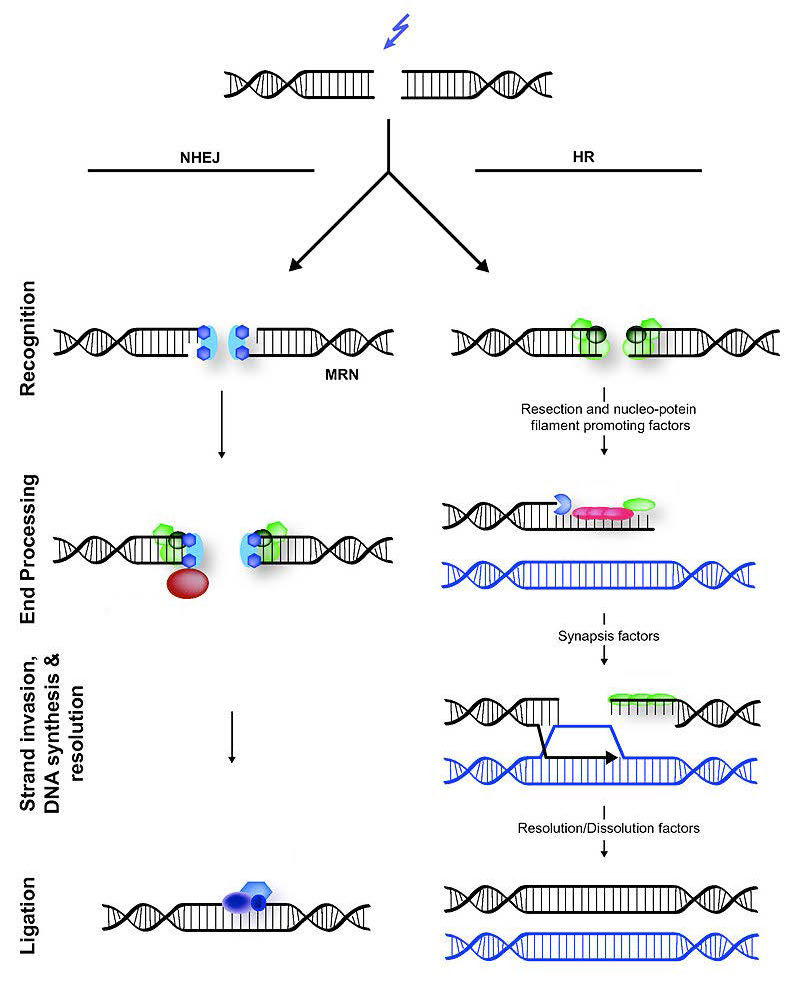
Figura 7.45 - Unión de extremos no homólogos (izquierda) versus recombinación homóloga (derecha) - Wikipedia
La reparación por recombinación homóloga ocurre comúnmente en las fases S y G2 tardías de la célula, cuando cada cromosoma ha sido replicado y la información de una cromátida hermana puede usarse como plantilla para lograr una reparación sin errores. Obsérvese que a diferencia de la reparación por escisión, donde se retiró la hebra dañada y la cadena hermana no dañada sirvió como molde para rellenar la región dañada, HR debe usar la información de otra molécula de ADN, ya que ambas cadenas del ADN están dañadas en las DSB.
Acción nucleasa
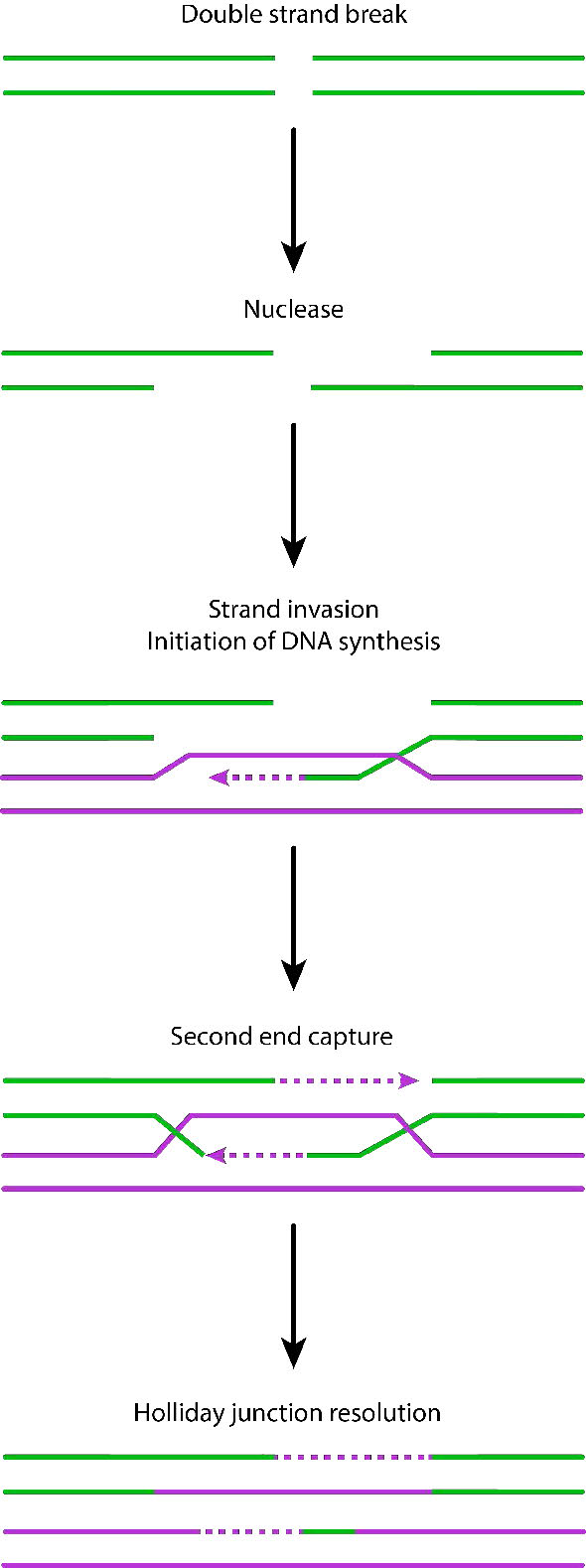
La detección de la rotura bicatenaria desencadena la actividad nucleasa que mastica una hebra en cada extremo de la rotura. Esto da como resultado la producción de salientes 3' monocatenarios en cada extremo. Estos extremos monocatenarios están unidos por varias proteínas, creando un filamento de nucleoproteína que luego puede “buscar” secuencias homólogas (coincidentes) en una cromátida hermana.
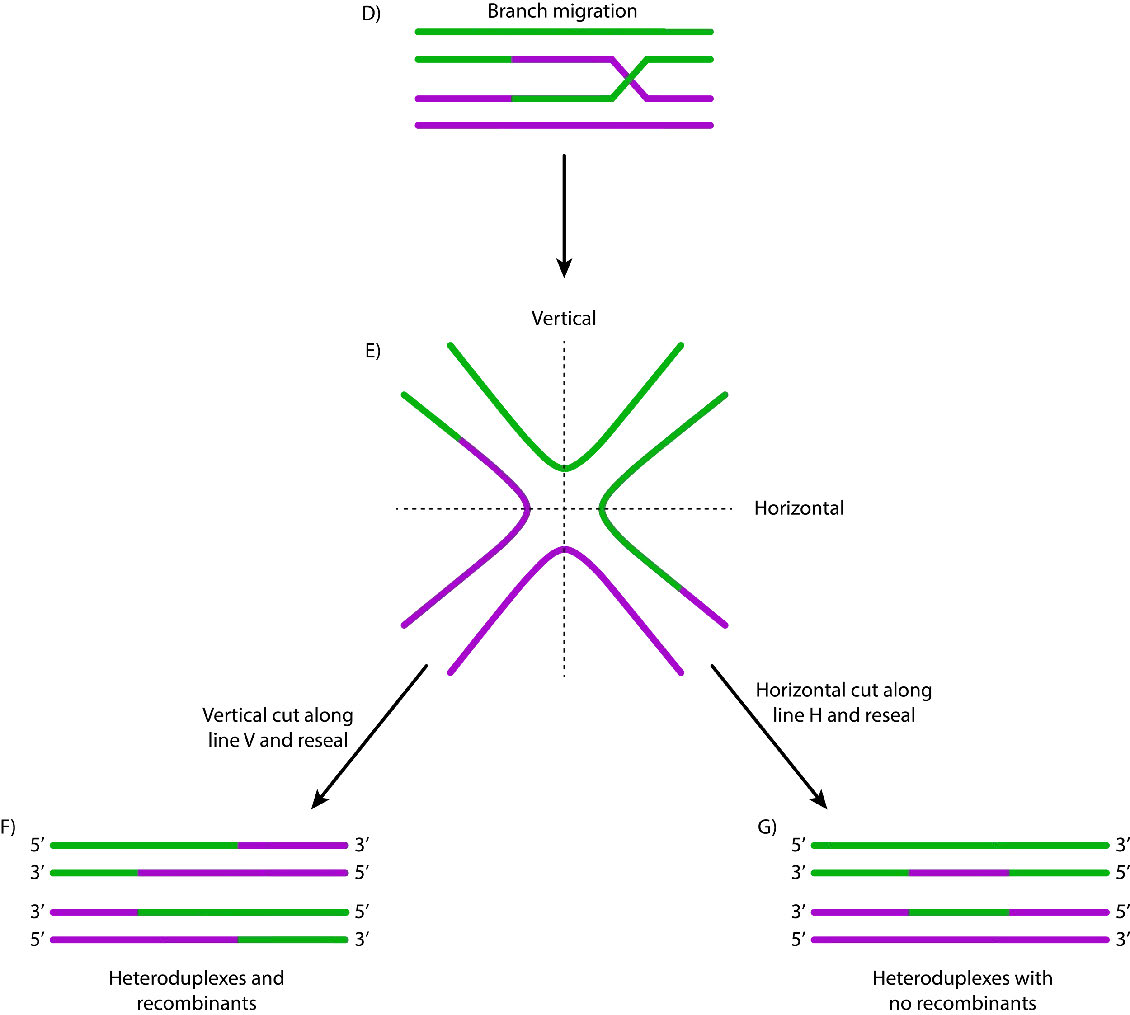
Cuando se encuentran tales secuencias, el filamento de nucleoproteína invade la cromátida hermana intacta, formando un cruce. Esto crea heterodúplex formados por hebras de ADN de diferentes cromátidas. La invasión de cadenas (Figura 7.47) es seguida por la migración de ramas, durante la cual la unión Holliday se mueve a lo largo del ADN, extendiendo el heterodúplex lejos del sitio original del cruce (Figura 7.48). En E. coli, la migración de ramas depende de la actividad de dos proteínas, RuVA y RuvB. El intermedio de recombinación resultante puede resolverse, con la ayuda de RuvC para dar hebras completas y libres de errores.
Unión de extremos no homólogos
A diferencia de la recombinación homóloga, la unión de extremos no homólogos (NHEJ) es propensa a errores. No utiliza ni requiere una plantilla homóloga para copiar, y funciona simplemente masticando los extremos rotos de los DSB y ligándolos juntos. No es sorprendente que NHEJ introduzca deleciones en el ADN como resultado.
Síntesis de ADN translesional
Como hemos visto, las células tienen una variedad de mecanismos para ayudar a salvaguardar la integridad de la información en el ADN. Una medida de último recurso es la síntesis de ADN translesional, también conocida como síntesis de bypass. La síntesis de translesión ocurre cuando una ADN polimerasa encuentra daño en el ADN en la cadena molde, pero en lugar de estancarse o saltarse más allá del daño, la replicación cambia a un modo propenso a errores, ignorando el molde e incorporando nucleótidos aleatorios en la nueva cadena. En E. coli, la síntesis de translesiones depende de las actividades de las proteínas codificadas por los genes uMuC y uMUD. Bajo las condiciones apropiadas (ver respuesta SOS, a continuación) uMuC y uMUD se activan para iniciar la síntesis de bypass. Siendo propensa a errores, la síntesis translesional da lugar a muchas mutaciones.
La respuesta SOS
Llamado así por las señales de socorro SOS estándar, el término “reparación SOS” se refiere a una respuesta celular al daño UV. Cuando las células bacterianas sufren daños extensos en su ADN como consecuencia de la exposición a los rayos UV, encienden la expresión coordinada de una gran cantidad de genes que son necesarios para la reparación del ADN. Estos incluyen los genes uvr necesarios para la reparación de la escisión de nucleótidos y recA, que participa en la recombinación homóloga. Además de estos mecanismos, que pueden llevar a cabo una reparación libre de errores, la respuesta SOS también puede inducir la expresión de polimerasas translesionadas codificadas por los genes dinA, dinB y umuCD.
¿Cómo se inducen todos estos genes de manera coordinada después del daño UV? Todos los genes inducidos en la respuesta SOS están regulados por dos componentes. El primero es la presencia de una secuencia de ADN corta aguas arriba de su región codificante, llamada caja SOS. La segunda es una proteína, el represor lexA (Figura 7.49), que se une a la caja SOS e impide la transcripción de los genes aguas abajo. La expresión de los genes requiere la eliminación de lexA de su sitio de unión. ¿Cómo se logra esto?
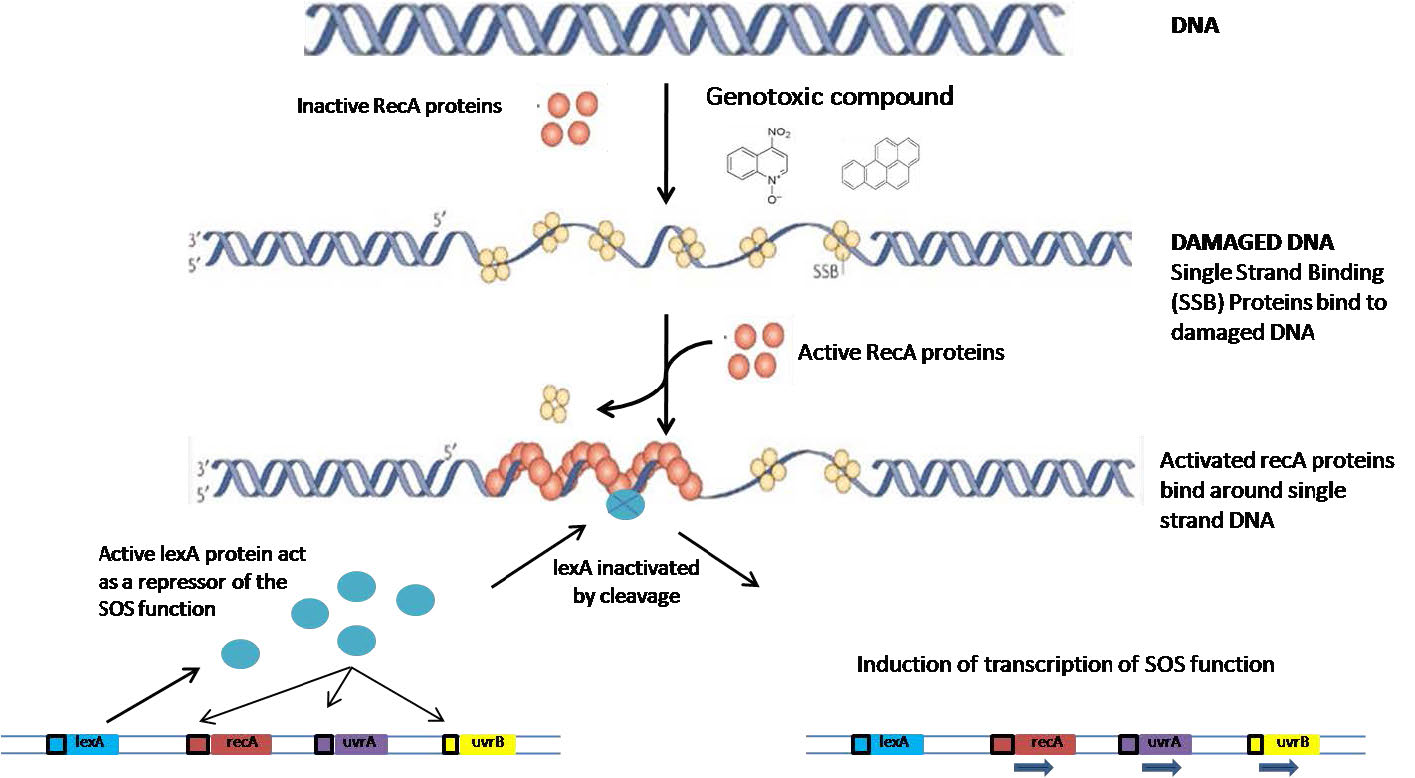
Cuando la exposición a la radiación produce roturas de ADN, la presencia de regiones monocatenarias desencadena la activación y unión de proteínas RecA a la región monocatenaria, creando un filamento de nucleoproteína. La interacción del recA con el represor lexA conduce a la autoescisión del represor, permitiendo que se expresen los genes aguas abajo (Figura 7.50).
Los genes controlados por el represor lexA, como se mencionó anteriormente, codifican proteínas que son necesarias para la reparación precisa del ADN, así como la síntesis de translesiones propensas a errores. Los diversos genes involucrados en la reparación del ADN son inducidos en un orden específico. En las etapas iniciales, los genes reparadores que son desreprimidos son para la reparación por escisión de nucleótidos, seguido de recombinación homóloga, ambos mecanismos de reparación libres de errores. Si el daño es demasiado extenso para ser reparado por estos sistemas, los mecanismos de reparación propensos a errores pueden ponerse en juego como último recurso.
Respuesta SOS y resistencia a antibióticos
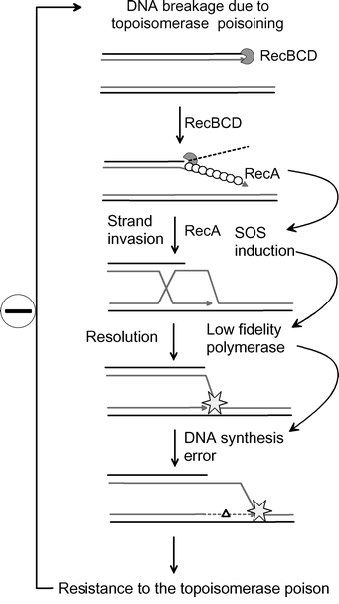
El aumento de la tasa de mutación en la respuesta SOS puede desempeñar un papel en la adquisición de resistencia a antibióticos en bacterias (Figura 7.51).
Un ejemplo es el desarrollo de resistencia a los venenos de topoisomerasas como la familia de fármacos de las fluoroquinolonas. Las fluoroquinolonas inhiben la capacidad de las topoisomerasas para religar los extremos de sus sustratos después de mellarlos para permitir que el ADN sobreenrollado se relaje. Esto da como resultado la acumulación de roturas de hebra que pueden desencadenar la respuesta SOS. Como consecuencia de la síntesis de ADN propensa a errores por polimerasas de baja fidelidad durante la respuesta SOS, hay un gran aumento en el número de mutaciones. Si bien algunas mutaciones pueden ser letales para la bacteria, otras pueden contribuir al rápido desarrollo de la farmacorresistencia en la población.