
8.1: Dinámica de dos especies que interactúan
- Page ID
- 52944

En la primera parte de este libro se han visto las dos categorías principales de dinámicas de una sola especie: logística y ortóloga, siendo el crecimiento exponencial una línea divisoria infinitamente fina entre ambas. Y ya has visto cómo la dinámica poblacional puede ser simple o caóticamente compleja.
En el futuro, verá tres tipos de dinámicas de dos especies: mutualismo, competencia y predación, y exactamente cuarenta tipos de dinámicas de tres especies, derivadas de los parámetros de las ecuaciones poblacionales y sus diversas combinaciones.
Para revisar, la dinámica poblacional de una sola especie se resume en la siguiente ecuación.
\[\frac{1}{N}\frac{dN}{dt}\,=\,r\,+\,sN\]
Aquí el parámetro r es la “tasa de crecimiento intrínseco” de la especie, la tasa neta a la que se introducen nuevos individuos a la población cuando la población es escasamente dispersa, y s es un parámetro de “dependencia de la densidad” que refleja cómo el tamaño de la población afecta al conjunto tasa. El parámetro s es clave. Si s es negativo, la población crece “logísticamente”, aumentando a una “capacidad de carga” de − r/s, o disminuyendo a esa capacidad de carga si la población comienza por encima de ella. Si s es positivo, entonces la población crece “ortologísticamente”, aumentando cada vez más rápido hasta encontrar algún límite no especificado no abordado en la ecuación. El crecimiento exponencial es la línea divisoria entre estos dos resultados, pero esto solo ocurriría si s permaneciera exactamente igual a cero.
¿Cómo debería extenderse esta ecuación de una sola especie a dos especies? Primero, en lugar de un número N para el tamaño poblacional de una especie, necesitamos un N para cada especie. Llame a estos N 1 para la especie 1 y N 2 para la especie 2. Entonces, si las dos especies no interactúan en absoluto, las ecuaciones podrían ser
\[\frac{1}{N_1}\frac{dN_1}{dt}\,=\,r_1\,+\,s_{1,1}\,N_1\]
\[\frac{1}{N_2}\frac{dN_2}{dt}\,=\,r_2\,+\,s_{2,2}\,N_2\]
Aquí r 1 y r 2 son las tasas de crecimiento intrínseco para N 1 y N 2, respectivamente, y s 1,1 y s 2,2 son los parámetros de dependencia de densidad para las dos especies. (Los subíndices emparejados en las ecuaciones de dos especies nos ayudan a abordar todas las interacciones).

Por lo tanto, hay cuatro posibles parámetros s i, j aquí:
- s 1,1: Cómo la densidad de la especie 1 afecta su propio crecimiento.
- s 1,2: Cómo la densidad de especies 2 afecta el crecimiento de la especie 1.
- s 2,1: Cómo la densidad de la especie 1 afecta el crecimiento de la especie 2.
- s 2,2: Cómo la densidad de la especie 2 afecta su propio crecimiento.
Con estos parámetros en mente, aquí están las ecuaciones de dos especies. Los nuevos términos de interacción están en azul a la derecha.
\[\frac{1}{N_1}\,\frac{dN_1}{dt}\,=\,r_1\,+\,s_{1,1}N_1\,+\,\color{blue}{s_{1,2}N_2}\]
\[\frac{1}{N_2}\,\frac{dN_2}{dt}\,=\,r_2\,+\,s_{2,2}N_2\,+\,\color{blue}{s_{2,1}N_1}\]
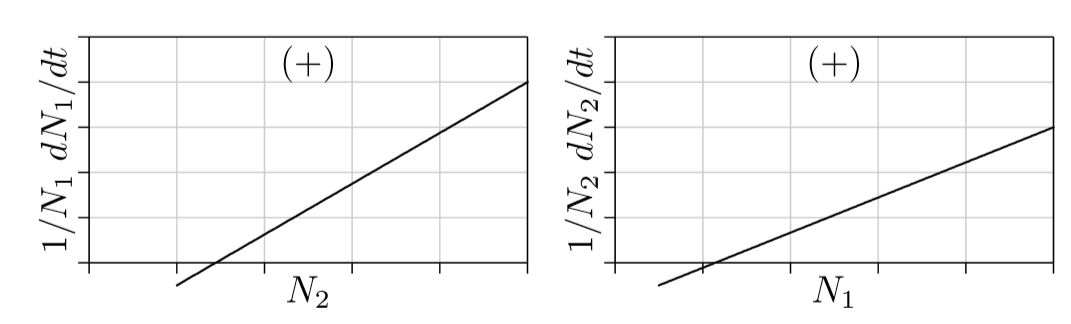
En las ecuaciones de una sola especie, el signo del término s separa los dos tipos principales de dinámica poblacional: positiva para ortóloga, negativa para logística. De igual manera, en las ecuaciones de dos especies, los signos de los parámetros de interacción s 1,2 y s 2,1 determinan la dinámica poblacional.
Dos parámetros permiten tres posibilidades principales: (1) ambos parámetros pueden ser negativos, (2) ambos pueden ser positivos, o (3) uno puede ser positivo y el otro negativo. Estas son las principales posibilidades con las que tiene que trabajar la selección natural.
Figura\(\PageIndex{1}\). Ambos parámetros de interacción negativos, competencia.

Competencia. Primero considere el caso donde s 1,2 y s 2,1 son ambos negativos, como en la Figura\(\PageIndex{1}\).
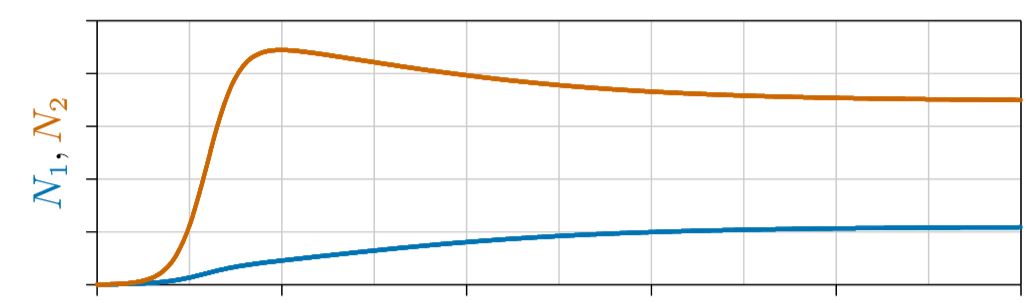
Para una sola especie, el parámetro s negativo hace que la población se acerque a una capacidad de carga. Lo mismo podría esperarse cuando los parámetros s 1,2 y s 2,1 son negativos: una o ambas especies se acercan a una capacidad de carga en la que la población permanece constante, o tan constante como lo permitan las condiciones ambientales externas.
Un ejemplo se muestra en la Figura\(\PageIndex{2}\), donde se traza la población de cada especie en el eje vertical y el tiempo en el eje horizontal. Aquí la Especie 2, en rojo, crece más rápido, gana la ventaja temprano y sube a un nivel alto. La especie 1, en azul, crece más lentamente pero eventualmente se eleva y, debido a la inhibición mutua entre especies en competencia, retrocede la población de la Especie 2. Las dos especies finalmente se acercan a una capacidad de carga conjunta.
En otros casos de competencia, un “competidor superior” puede llevar al otro competidor a la extinción, un resultado llamado “exclusión competitiva”. O bien, cualquiera de las especies puede llevar a la otra a la extinción, dependiendo de cuál gane la ventaja primero. Estos y otros casos se tratan en capítulos posteriores.
En cualquier caso, cuando ambos términos de interacción s 1,2 y s 2,1 son negativos, en interacción minús—menos, cada especie inhibe el crecimiento de la otra, lo que los ecologistas llaman la “competencia de interacción”.
Mutualismo. Lo contrario de la competencia es el mutualismo, donde cada especie potencia más que inhibe el crecimiento de la otra. Tanto s 1,2 como s 2,1 son positivos.
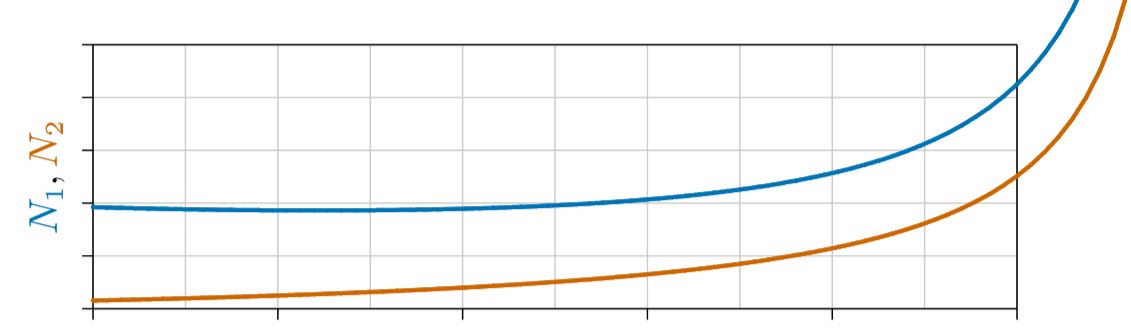
En la Figura\(\PageIndex{3}\) se representa una forma de “mutualismo obligado”, donde ambas especies declinan a la extinción si alguna de ellas no está presente. Esto es análogo a un punto conjunto de Allee, donde las curvas de crecimiento cruzan el eje horizontal y se vuelven negativas por debajo de ciertos niveles críticos de población. Si no es así y las curvas de crecimiento cruzan el eje vertical, cada especie puede sobrevivir sola; esto se llama “mutualismo facultativo”, y aprenderemos más sobre ello en capítulos posteriores.
Por ahora, el punto importante es cómo las poblaciones mutualistas crecen o disminuyen con el tiempo. Una sola especie cuya densidad de alguna manera mejora su propio ritmo de crecimiento se vuelve ortóloga, aumentando cada vez más rápidamente hacia una singularidad, ante la cual crecerá tan numerosa que será comprobada por algún otro límite inevitable, como el espacio, la depredación o la enfermedad.
Resulta que la dinámica de dos especies potenciando el crecimiento de la otra es similar a la de una sola especie potenciando su propio crecimiento. Ambos se mueven a una singularidad a tasas cada vez mayores, como se ilustra anteriormente en la Figura 4.2.1 y abajo en la Figura\(\PageIndex{4}\). Desde luego, tal crecimiento no puede continuar para siempre. Eventualmente será comprobada por alguna fuerza más allá del alcance de las ecuaciones, así como el crecimiento de la población humana se comprobó abruptamente a mediados del siglo XX, tan claramente visible antes en la Figura 6.3.1.
Depredación. La posibilidad restante para estas ecuaciones de dos especies es cuando un parámetro de interacción si, j es positivo y el otro es negativo. Es decir, cuando la primera especie potencia el crecimiento de la segunda mientras que la segunda especie inhibe el crecimiento de la primera. O viceversa. Esto es “depredación”, que también se manifiesta como parasitismo, enfermedad y otras formas.
Piensa en un depredador y en su presa. Cuantas más presas, más fácil les resulta a los depredadores atraparlas, de ahí que más fácil sea para los depredadores alimentar a sus crías y mayor será el crecimiento poblacional del depredador. Esta es la parte derecha de la Figura\(\PageIndex{5}\). Cuantos más depredadores haya, sin embargo, más presas son capturadas; de ahí que menor sea la tasa de crecimiento de la presa, como se muestra a la izquierda de la figura. N 1 aquí, entonces, representa a la presa, y N 2 representa al depredador.
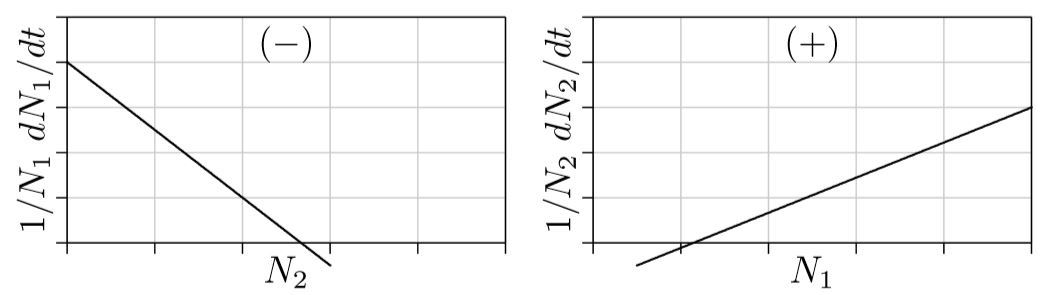
Las presas pueden sobrevivir solas, sin depredadores, como se refleja a la izquierda en crecimiento positivo para N 1 cuando N 2 es 0. Los depredadores, sin embargo, no pueden sobrevivir sin presas, como se refleja a la derecha en el crecimiento negativo para N 2 cuando N 1 es 0. Esto es como un punto Allee para los depredadores, que empezará a morir si la población de presas cae por debajo de este punto.
La pregunta aquí es esta: ¿cuál será la dinámica poblacional de depredador y presa a través del tiempo? ¿Las poblaciones crecerán logísticamente y se nivelarán en estado estacionario, como sugiere el parámetro negativo s 1,2, o aumentarán ortologísticamente, como sugiere el parámetro positivo s 2,1?
En realidad, hacen ambas cosas. A veces aumentan más rápido que exponencialmente, cuando las poblaciones de depredadores son bajas y las poblaciones de presas en crecimiento proporcionan tasas de crecimiento per cápita cada vez mayores para el depredador, según la parte derecha de la Figura\(\PageIndex{5}\). Sin embargo, a su debido tiempo, los depredadores se vuelven abundantes y deprimen a las poblaciones de presas, lo que a su vez reduce el crecimiento de las Como se muestra en la Figura\(\PageIndex{6}\), las poblaciones oscilan en tensiones continuas entre depredador (línea roja) y presa (línea azul).
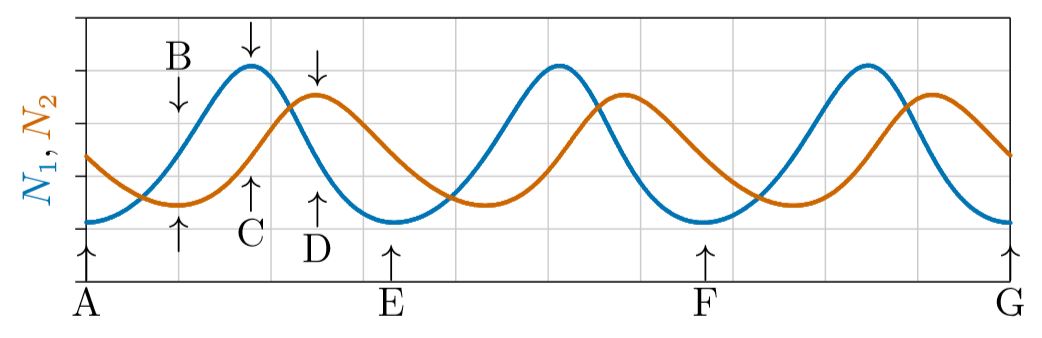
Examina esta figura en detalle. Al inicio, etiquetada como A, la población de presas es baja y los depredadores están disminuyendo por falta de alimento. Una disminución constante en el número de depredadores crea mejores y mejores condiciones para las presas, cuyas poblaciones luego aumentan ortologísticamente a tasas per cápita cada vez más aceleradas a medida que los depredadores mueren y las condiciones para la presa mejoran en consecuencia.
Pero entonces la situación gira. Las presas crecen abundantes, con la población elevándose por encima del punto Allee del depredador, en B. Así, el número de depredadores comienza a aumentar. Si bien las poblaciones de depredadores son bajas y el número de presas aumenta, las condiciones mejoran continuamente para los depredadores, y sus poblaciones crecen aproximadamente ortólogicamente por un tiempo.
Entonces los depredadores se vuelven abundantes e impulsan la tasa de crecimiento de la presa negativa. La situación vuelve a cambiar, en C. Prey comienzan a disminuir y el crecimiento de los depredadores se vuelve aproximadamente logístico, nivelándose y comenzando a disminuir en D. Por E ha llegado al círculo completo y el proceso se repite, ad infinitum.
Mientras que la Figura\(\PageIndex{6}\) ilustra la forma clásica para las interacciones depredador-presa, otras formas son posibles. Cuando las condiciones son adecuadas, las oscilaciones pueden amortiguar y tanto las poblaciones de depredadores como presas pueden alcanzar estados estables. O las oscilaciones pueden llegar a ser tan salvajes que los depredadores matan a todas las presas y luego se desvanecen ellos mismos. Esto supone algún valor efectivo de cero para N 1 y N 2, por debajo del cual se “ajustan” a cero. O las poblaciones de presas pueden llegar a ser tan bajas que todos los depredadores mueren, dejando a la presa en paz. O ambos pueden extinguirse. O, en el caso de los depredadores humanos, la presa puede ser domesticada y transformada en mutualistas. Más sobre todas esas dinámicas en capítulos posteriores.