
12.2: Regulación génica en procariotas
- Page ID
- 54489

Muchos genes procariotas están organizados en operones, genes enlazados transcritos en un solo ARNm que codifica dos o más proteínas. Los operones suelen codificar proteínas con funciones relacionadas. La regulación de la actividad de un operón (en lugar de múltiples genes individuales que codifican proteínas individuales) permite una mejor coordinación de la síntesis de varias proteínas a la vez. En E. coli, el operón lac regulado codifica tres enzimas involucradas en el metabolismo de la lactosa (un nutriente alternativo a la glucosa). La regulación de un operón (o de un solo gen para el caso) puede ser por represión o por inducción. Cuando un metabolito pequeño en una célula se une a una proteína represora o inductora reguladora, la proteína sufre un cambio alostérico que le permite unirse a una secuencia reguladora de ADN..., o desunirse del ADN. Veremos ejemplos de dicha regulación en los operones lac y trp. La regulación génica del operón Lac es un ejemplo de represión génica así como de inducción. La regulación del operón Trp (triptófano) es por represión génica. En ambos operones, los cambios en los niveles de metabolitos intracelulares reflejan el estado metabólico de la célula y provocan cambios apropiados en la transcripción génica. Veremos la regulación de ambos operones.
216 Descripción general de la regulación de genes procariotas
El ARNm transcrito del operón lac se traduce simultáneamente en esas tres enzimas, como se muestra a continuación.
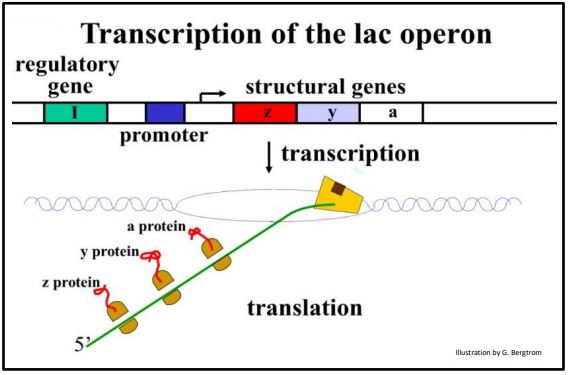
A. Mecanismos de Control del Operón Lac
En el tracto digestivo animal (incluido el nuestro), los genes del operón lac de E. coli regulan el uso de lactosa como nutriente alternativo a la glucosa. ¡Piensa en queso en lugar de chocolate! El operón consiste en genes lacZ, lacY y lacA que se denominaron genes estructurales. Por definición, los genes estructurales codifican proteínas que participan en la estructura celular y la función metabólica. Como ya se señaló, el operón lac se transcribe en un ARNm que codifica las proteínas Z, Y y A.
Echemos un vistazo más de cerca a la estructura del operón lac y la función de las proteínas Y, Z y A (abajo).
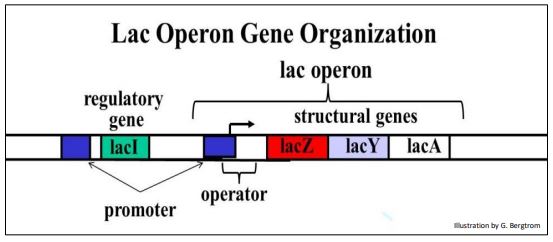
El gen lacZ codifica β-galactosidasa, la enzima que rompe la lactosa (un disacárido) en galactosa y glucosa. El gen lacY codifica la lactosa permeasa, una proteína de membrana que facilita la entrada de lactosa en las células. No se entiende bien el papel del gen lacA (una transacetilasa) en el metabolismo energético de la lactosa. El gen I a la izquierda del gen lac Z es un gen regulador (para distinguirlo de los genes estructurales). Los genes reguladores codifican proteínas que interactúan con secuencias de ADN reguladoras asociadas con un gen para controlar la transcripción. La secuencia operadora que separa los genes I y Z es una secuencia de ADN reguladora de la transcripción.
El operón lac de E. coli suele ser silencioso (reprimido) porque estas células prefieren la glucosa como fuente de energía y carbono. En presencia de suficiente glucosa, una proteína represora (el producto del gen I) se une al operador, bloqueando la transcripción del operón lac. Incluso si se dispone de lactosa, las células no la usarán como fuente alternativa de energía y carbono cuando los niveles de glucosa sean adecuados. Sin embargo, cuando los niveles de glucosa bajan, el operón lac está activo y los tres productos enzimáticos se traducen. Veremos cómo los niveles limitantes de glucosa inducen la transcripción máxima del operón lac tanto por desrepresión como por inducción directa, conduciendo a la transcripción máxima de los genes lac solo cuando sea necesario (es decir, en presencia de lactosa y ausencia de glucosa). Veamos algunos de los experimentos clásicos que nos llevaron a comprender la regulación del gen de E. coli en general, y del operón lac en particular.
A finales de la década de 1950 y principios de los sesenta, Francois Jacob y Jacques Monod estudiaban el uso de diferentes azúcares como fuentes de carbono por E. coli. Sabían que E. coli de tipo silvestre no produciría las proteínas\(\beta \) -galactosidasa,\(\beta \) -galactósido permeasa o\(\beta \) -galactósido transacetilasa cuando se cultivaban en glucosa. ¡Por supuesto, también sabían que las células cambiarían a lactosa para su crecimiento y reproducción si se les privaba de glucosa! Luego buscaron y aislaron diferentes mutantes de E. coli que no podían crecer con lactosa, incluso cuando no había glucosa en el medio de crecimiento. Estos son algunos de los mutantes que estudiaron:
- Un mutante no logró producir enzima\(\beta \) -galactosidasa activa pero produjo permeasa.
- Un mutante no logró producir permeasa activa pero produjo cantidades normales de\(\beta \) -galactosidasa.
- Otro mutante no logró producir transacetilasa..., pero aún podría metabolizar la lactosa en ausencia de glucosa. De ahí la incertidumbre de su papel en el metabolismo de la lactosa.
- Curiosamente, ¡una cepa mutante no logró producir ninguna de las tres enzimas!
Dado que los mutantes dobles son muy raros y los mutantes triples aún más raros, Jacob y Monod inferían que la activación de los tres genes en presencia de lactosa se controlaron juntos de alguna manera. De hecho, fue este descubrimiento el que definió al operón como un conjunto de genes transcritos como un solo ARNm, cuya expresión podría por lo tanto coordinarse de manera efectiva. Posteriormente caracterizaron la proteína represora producida por el gen lacI. Jacob, Monod y Andre Lwoff compartieron el Premio Nobel de Medicina en 1965 por su trabajo sobre la regulación de genes bacterianos. Ahora sabemos que la regulación negativa y positiva del operón lac (descrita a continuación) dependen de dos proteínas reguladoras que juntas, controlan la tasa de metabolismo de la lactosa.
1. Regulación Negativa del Operón Lac por Lactosa
Consulte la siguiente ilustración para identificar a los jugadores en la desrepresión del operón lac.
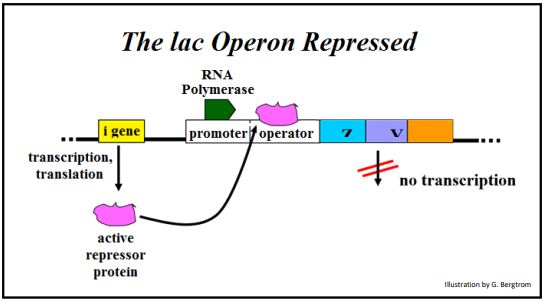
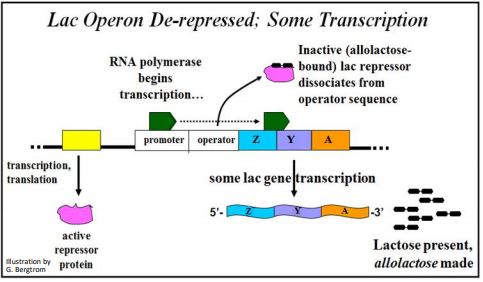
El producto proteico represor del gen I siempre se elabora y está presente en las células de E. coli. ¡La expresión del gen I no está regulada! En ausencia de lactosa en el medio de crecimiento, la proteína represora se une fuertemente al ADN del operador. Mientras que la ARN polimerasa está unida al promotor y lista para transcribir el operón, la presencia del represor unido a la secuencia operadora cercana al gen Z bloquea físicamente su movimiento hacia adelante. Bajo estas condiciones, se realiza poca o ninguna transcripción. Si las células se cultivan en presencia de lactosa, la lactosa que ingresa a las células se convierte en alolactosa. La alolactosa se une al represor sentado en el ADN del operador para formar un complejo de 2 partes, como se muestra a continuación.
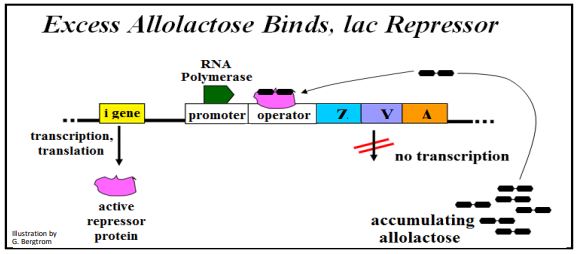
El represor alterado alostéricamente se disocia del operador y la ARN polimerasa puede transcribir los genes del operón lac como se ilustra a continuación.
2. Regulación Positiva del Operón Lac; Inducción por Activación de Catabolitos
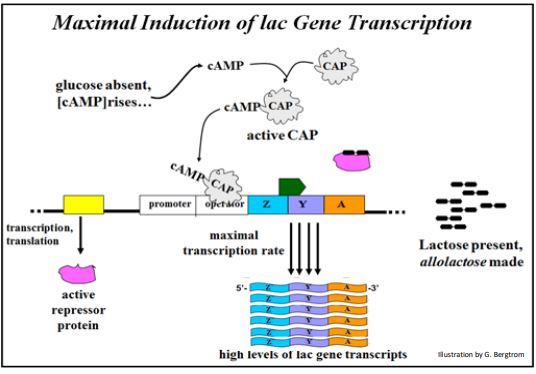
El segundo mecanismo de control que regula la expresión del operón lac está mediado por CAP (proteína activadora de catabolito unida a AMPc o proteína receptora de cAMP) Cuando hay glucosa disponible, los niveles celulares de AMPc son bajos en las células y la CAP está en una conformación inactiva. Por otro lado, si los niveles de glucosa son bajos, los niveles de AMPc suben y se unen a la PAC, activándola. Si los niveles de lactosa también son bajos, el CAP unido a AMPc no tendrá ningún efecto. Si hay lactosa y los niveles de glucosa son bajos, entonces la alolactosa se une al represor lac provocando que se disocie de la región operadora. Bajo estas condiciones, la CAP unida a cAMP puede unirse al operador en lugar de la proteína represora. En este caso, en lugar de bloquear la ARN polimerasa, el CAP unido a cAMP activado induce una transcripción del operón lac aún más eficiente. El resultado es la síntesis de mayores niveles de enzimas lac que facilitan el uso celular eficiente de la lactosa como alternativa a la glucosa como fuente de energía. La activación máxima del operón lac en lactosa alta y glucosa baja se muestra a continuación.
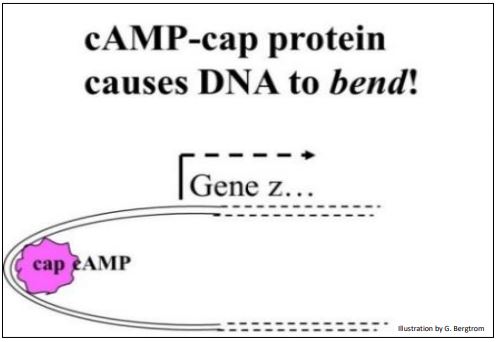
La CAP unida a cAMP es un inductor de transcripción. Lo hace forzando a doblar el ADN en la región promotor-operador. Y dado que doblar la doble hélice afloja los enlaces H, se vuelve más fácil para la ARN polimerasa encontrar y unir el promotor en la cadena de ADN a transcribir..., y para que comience la transcripción La flexión del ADN inducida por cAMP-CAPA se ilustra a continuación.
3. Regulación del operón Lac por exclusión de inductores y múltiples operadores
En los últimos años, se han descubierto capas adicionales de regulación del operón lac. En un caso, se regula la capacidad de la permeasa lac para transportar lactosa a través de la membrana celular. En otra, se ha descubierto que secuencias operadoras adicionales interactúan con un represor multimérico para controlar la expresión del gen lac.
a) Regulación del uso de Lactosa por Exclusión Inductora
Cuando los niveles de glucosa son altos (incluso en presencia de lactosa), se consume fosfato para fosforilar intermedios glicolíticos, manteniendo bajos los niveles de fosfato citoplásmico. En estas condiciones, elIIaGLC no fosforilada se une a la enzima lactosa permeasa en la membrana celular, evitando que lleve lactosa a la célula.
A continuación se muestra el papel de EIIA Glc fosforilada y no fosforilada en la regulación del operón lac.
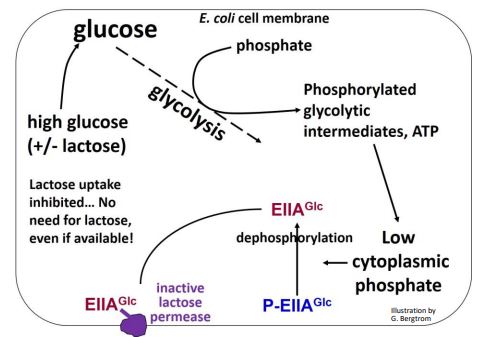
Los niveles altos de glucosa bloquean la entrada de lactosa en las células, previniendo eficazmente la formación de alolactosa y la desrepresión del operón lac. La exclusión inductora es así una forma lógica para que las células manejen una abundancia de glucosa, esté o no presente lactosa. Por otro lado, si los niveles de glucosa son bajos en el medio de crecimiento, las concentraciones de fosfato en las células aumentan lo suficiente para que una quinasa específica fosforile el EIIaGLC. EIIaGLC fosforilada sufre entonces un cambio alostérico y se disocia de la permeasa de lactosa, haciéndola activa para que más lactosa pueda ingresar a la célula. En otras palabras, ¡el inductor no está “excluido” bajo estas condiciones!
La quinasa que fosforila EIIA Glc es parte de una cascada del sistema fosfoenolpiruvato (PEP) - dependiente de fosfotransferasa (PTS). Cuando los niveles de glucosa extracelular son bajos, la célula activa el sistema PTS en un esfuerzo por traer cualquier glucosa que esté alrededor a la célula. Pero la última enzima en la cascada de fosforilación de PTS es la quinasa que fosforila EIIA Glc. La EIIA Glc fosforilada se disocia de la permeasa de lactosa, reactivándola, trayendo lactosa disponible a la célula desde el medio.
b) Estructura de Proteína Represora y Secuencias Operadoras Adicionales
El represor lac es un tetrámero de subunidades idénticas (abajo).
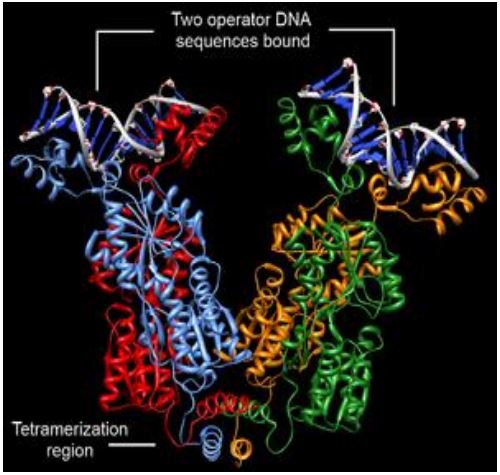
Cada subunidad contiene un motivo hélice-vuelta-hélice capaz de unirse al ADN. Sin embargo, la secuencia de ADN operadora aguas abajo del promotor en el operón consiste en un par de repeticiones invertidas separadas de tal manera que solo pueden interactuar con dos de las subunidades represivas, dejando desconocida la función de las otras dos subunidades... es decir, ¡hasta hace poco!
Dos regiones operadoras más se caracterizaron recientemente en el operón lac. Uno, llamado O 2, está dentro del gen lac z mismo y el otro, llamado O 3, se encuentra cerca del final de, pero dentro del gen lac I. Aparte de su ubicación inusual dentro de genes reales, estos operadores, que interactúan con las dos subunidades represivas restantes, no fueron detectados al principio porque las mutaciones en la región O2 o O3 individualmente no contribuyen sustancialmente al efecto de la lactosa en la desrepresión del operón lac. Solo mutar ambas regiones al mismo tiempo da como resultado una reducción sustancial en la unión del represor al operón.
B. Mecanismo de Control del Operón Triptófano
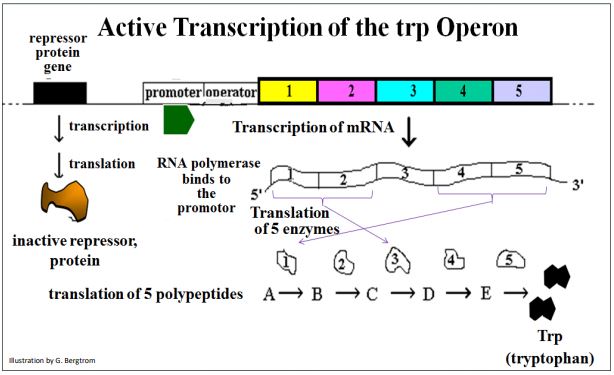
Si se dispone de un amplio triptófano (trp), la vía de síntesis de triptófano se puede inhibir de dos maneras. Primero, recuerde cómo la inhibición por retroalimentación por el exceso de trp puede inhibir alostéricamente la vía de síntesis de trp. Una respuesta rápida ocurre cuando el triptófano está presente en exceso, lo que resulta en una rápida inhibición de la retroalimentación al bloquear la primera de cinco enzimas en la vía de síntesis de trp. El operón trp codifica polipéptidos que componen dos de estas enzimas.
La enzima 1 es una proteína multimérica, elaborada a partir de polipéptidos codificados por los genes trp5 y trp4. Los productos de los genes trp1 y trp2 conforman la Enzima 3. Si los niveles celulares de triptófano bajan debido a que el aminoácido se consume rápidamente (por ejemplo, debido a la demanda de proteínas durante el crecimiento rápido), las células de E. coli continuarán sintetizando el aminoácido, como se ilustra a continuación.
Por otro lado, si el consumo de triptófano se ralentiza, el triptófano se acumula en el citoplasma. El exceso de triptófano se unirá al represor trp. El represor unido a trp luego se une al operador trp, bloqueando la ARN polimerasa para que no transcriba el operón. A continuación se muestra la represión del operón trp por trp.
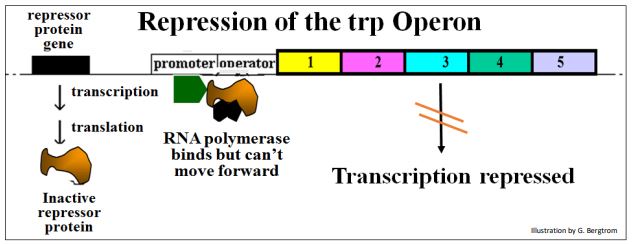
En este escenario, el triptófano es un co-represor. La función de un co-represor es unirse a una proteína represora y cambiar su conformación para que pueda unirse al operador.