
1.4: Lo que podemos (y no podemos) aprender sobre la historia evolutiva de las especies vivas
- Page ID
- 53944

Tradicionalmente, los científicos han utilizado fósiles para cuantificar tasas y patrones de evolución a través de largos períodos de tiempo (a veces llamados “macroevolución”). Estos enfoques han sido tremendamente informativos. Ahora tenemos una imagen detallada de la dinámica evolutiva de muchos grupos, desde homínidos hasta cocodrilos. En algunos casos, registros fósiles muy detallados de algunos tipos de organismos —por ejemplo, invertebrados marinos— han permitido realizar pruebas cuantitativas de modelos evolutivos particulares.
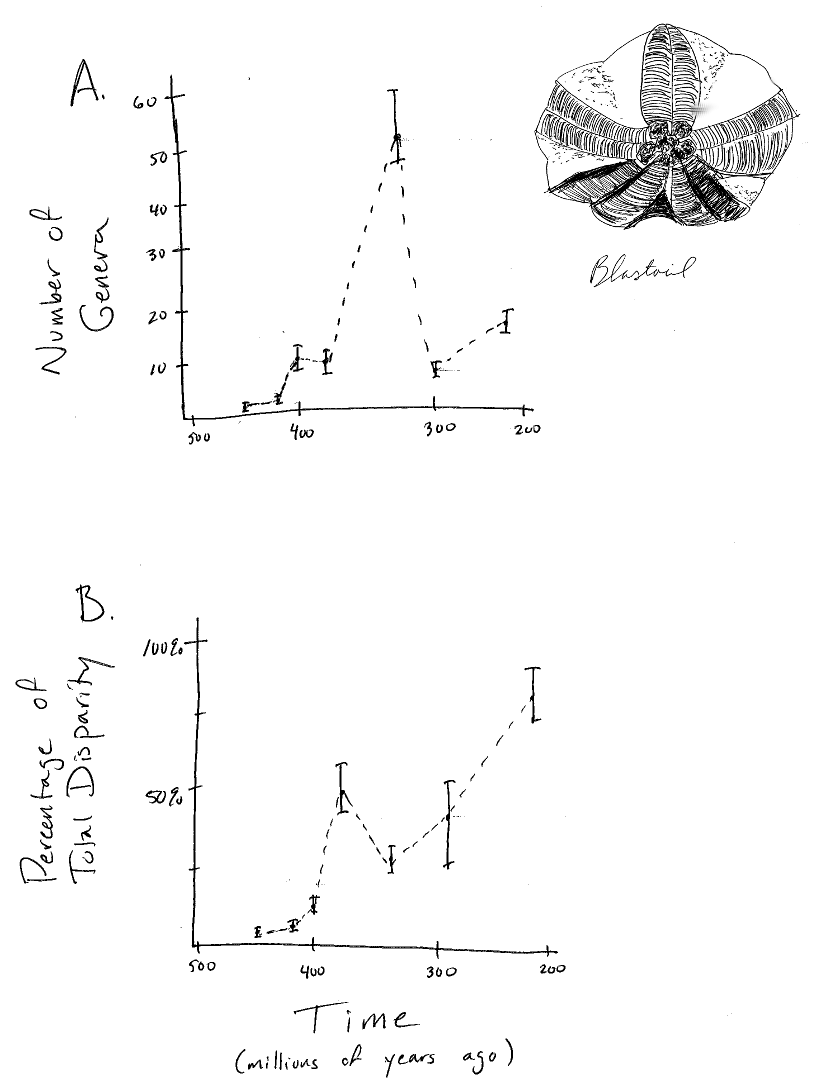
Los fósiles son particularmente buenos para mostrar cómo la diversidad de especies y los caracteres morfológicos cambian a través del tiempo. Por ejemplo, si uno tiene una secuencia de fósiles con tiempos conocidos de ocurrencia, se pueden reconstruir patrones de diversidad de especies a través del tiempo. Un ejemplo clásico de esto es la reconstrucción de Sepkoski (1984) de la diversidad de invertebrados marinos en los últimos 600 millones de años. También se pueden cuantificar los rasgos de esos fósiles y medir cómo cambian a través de varios intervalos de tiempo (por ejemplo, Foote 1997). En algunos grupos, podemos realizar parcelas de cambios en linaje y diversidad de rasgos simultáneamente (Figura 1.2). Los fósiles son la única evidencia que tenemos de linajes evolutivos que se han extinguido, y proporcionan valiosa evidencia directa sobre la dinámica evolutiva en el pasado.
Sin embargo, los enfoques basados en fósiles enfrentan algunos desafíos. La primera es que el registro fósil está incompleto. Se trata de un fenómeno bien conocido, identificado por el propio Darwin (aunque se siguen encontrando muchos fósiles nuevos). El registro fósil está incompleto de algunas formas muy particulares que a veces pueden obstaculizar nuestra capacidad de estudiar procesos evolutivos utilizando solo fósiles. Un ejemplo es que los fósiles son raros o están ausentes de algunos ejemplos clásicos de radiación adaptativa en las islas. Por ejemplo, todo el registro fósil de anoles caribeños, una conocida radiación adaptativa de lagartos, consiste en menos de diez especímenes conservados en ámbar (Losos 2009). De manera similar, carecemos de fósiles para otras radiaciones adaptativas como los cíclidos africanos y los pinzones de Darwin. La ausencia de fósiles en estos grupos limita nuestra capacidad para estudiar directamente las primeras etapas de la radiación adaptativa. Otra limitación del registro fósil se relaciona con especies y especiaciones. Es muy difícil identificar y clasificar especies en el registro fósil —incluso más difícil de lo que es hacerlo para las especies vivas. Es difícil distinguir a las especies, y particularmente difícil precisar el momento exacto en que las nuevas especies se separan de sus parientes cercanos. De hecho, la mayoría de los estudios de diversidad fósil se centran en grupos taxonómicos superiores como géneros, familias u órdenes (ver, por ejemplo, Sepkoski 1984). Estos estudios han sido inmensamente valiosos pero puede ser difícil conectar estos resultados con lo que sabemos sobre las especies vivas. De hecho, son las especies (y no géneros, familias u órdenes) las que forman las unidades básicas de casi todos los estudios evolutivos. Entonces, los fósiles tienen un gran valor pero también sufren algunas limitaciones particulares.
Los árboles filogenéticos representan una rica fuente de información complementaria sobre la dinámica de la formación de especies a través del tiempo. Los enfoques filogenéticos proporcionan un complemento útil a los fósiles porque sus limitaciones son muy diferentes de las limitaciones del registro fósil. Por ejemplo, a menudo se pueden incluir todas las especies vivas en un grupo al crear un árbol filogenético. Adicionalmente, se puede usar información de estudios detallados sistemáticos y taxonómicos para identificar especies, en lugar de enfrentar la ambigüedad inherente al usar fósiles. Los árboles filogenéticos proporcionan una fuente distinta de información sobre el cambio evolutivo que es complementaria a los enfoques basados en fósiles. Sin embargo, los árboles filogenéticos no proporcionan todas las respuestas. En particular, hay ciertos problemas que los datos comparativos por sí solos simplemente no pueden abordar. Los más destacados de estos, a los que volveré más adelante, son la reconstrucción de rasgos de ancestros particulares (reconstrucción del estado ancestral; Losos 2011) y distinguir entre ciertos tipos de modelos donde el tempo de la evolución cambia a través del tiempo (Slater et al. 2012). Algunos autores han argumentado que la extinción, también, no se puede detectar en forma de árbol filogenético (Rabosky 2010). Voy a argumentar en contra de este punto de vista en el Capítulo 11, pero la extinción sigue siendo un problema complicado cuando se limita a muestras de un solo intervalo de tiempo (la actualidad). Los árboles filogenéticos proporcionan una rica fuente de información sobre el pasado, pero debemos ser conscientes de sus limitaciones (Alroy 1999).
Quizás el mejor enfoque combinaría datos fósiles y filogenéticos directamente. Los paleontólogos que estudian fósiles y los neontólogos que estudian árboles filogenéticos comparten un conjunto común de modelos matemáticos. Esto significa que, en algún momento, los dos campos pueden fusionarse, y ambos tipos de información pueden combinarse para estudiar patrones evolutivos de manera cohesiva e integradora. Sin embargo, sorprendentemente, hasta el momento se ha realizado poco trabajo en esta área (pero ver Slater et al. 2012, Heath et al. (2014)).