
10.3: Detalles de la Transcripción
- Page ID
- 54161

Encuentre un resumen bien escrito de la transcripción en procariotas y eucariotas en un sitio web de los NIH (Transcripción en procariotas y eucariotas). Aquí (y en este enlace), encontrarás proteínas que se unen al ADN. Algunas proteínas se unen al ADN para regular la transcripción, induciendo o silenciando la transcripción de un gen. Discutiremos su papel en la regulación de la expresión génica más adelante. Otras proteínas interactúan con el ADN simplemente para permitir la transcripción. Estos incluyen uno o más que, junto con la propia ARN polimerasa, que deben unirse al promotor génico para iniciar la transcripción. Primero veremos la transcripción bacteriana.
A. Transcripción en Procariotas
En E. coli, una sola ARN polimerasa transcribe todo tipo de ARN, asociándose con una de varias proteínas del factor sigma (\(\sigma \)-factores) para iniciar la transcripción. Resulta que diferentes secuencias promotoras y\(\sigma \) los factores correspondientes juegan un papel en la transcripción de diferentes genes (ilustrado a continuación).
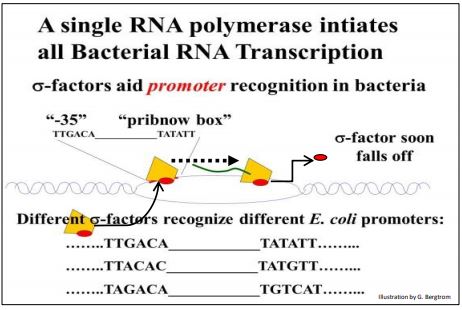
En ausencia del\(\sigma \) factor -la ARN polimerasa de E. coli puede transcribir ARN, pero lo hace a alta velocidad, y a partir de secuencias aleatorias en el cromosoma. En contraste, cuando el\(\sigma \) factor -se une a la ARN polimerasa, el complejo parece escanear el ADN, reconocer y luego unirse a la secuencia promotora de un gen. En este caso, la tasa de transcripción general es más lenta, pero solo se transcriben genes, ¡en lugar de bits aleatorios del genoma bacteriano! La caja Pribnow, llamada así por su descubridor, fue la primera secuencia promotora caracterizada.
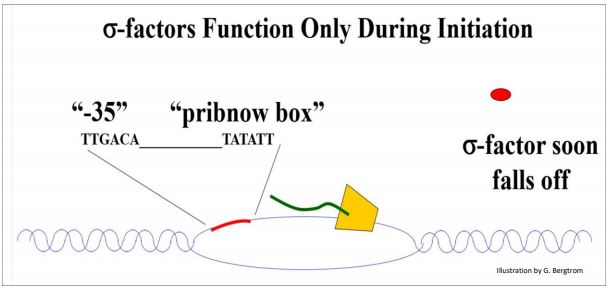
La elongación es la adición sucesiva de nucleótidos complementarios a sus moldes de ADN, formando enlaces fosfodiéster. Las reacciones enzimáticas de elongación son similares a la elongación catalizada por ADN polimerasa durante la replicación.
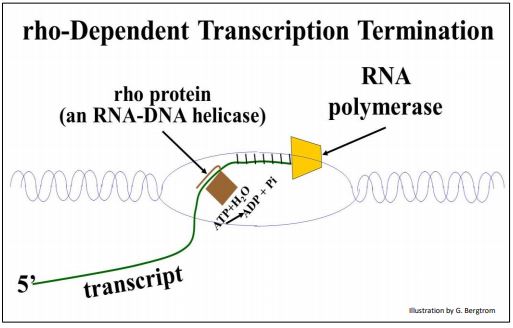
Hay dos formas en que la ARN polimerasa bacteriana 'sabe' cuándo ha llegado al final de una unidad de transcripción. En un caso, a medida que la ARN polimerasa se acerca al extremo 3' del transcrito naciente, transcribe una región rica en C de 72 bases. En este punto, un factor de terminación llamado la proteína rho se une a la cadena de ARN naciente. Rho es una helicasa dependiente de ATP que rompe los enlaces H entre el ARN y la cadena de ADN molde, evitando así una mayor transcripción.
La terminación dependiente de Rho se ilustra a continuación.
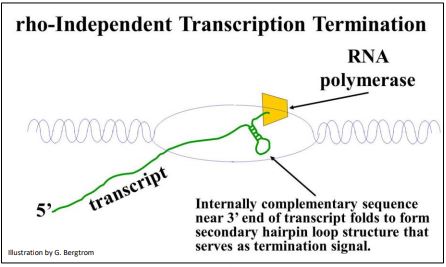
En el otro mecanismo de terminación, la polimerasa transcribe ARN cuya señal de terminación asume una estructura de bucle de horquilla secundaria que provoca la disociación de la ARN polimerasa, ADN molde y el nuevo transcrito de ARN. A continuación se ilustra el papel del bucle de horquilla en la terminación independiente de rho-independent.
B. Transcripción en Eucariotas
Mientras que las bacterias dependen de una sola ARN polimerasa para sus necesidades de transcripción, los eucariotas utilizan tres ARN polimerasas diferentes para sintetizar los tres tipos principales de ARN diferentes, como se muestra a continuación.
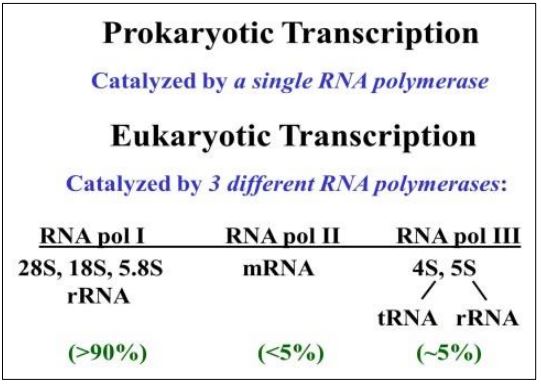
Nótese que la catálisis de la síntesis de la mayor parte del ARN en una célula eucariota (ARNr) es por ARN polimerasa I. Con la ayuda de proteínas de iniciación, cada ARN polimerasa inicia la transcripción en una secuencia promotora. Una vez iniciadas, las ARN polimerasas catalizan la formación sucesiva de enlaces fosfodiéster para alargar el transcrito. Recordemos que los ARNm son los menos abundantes en eucariotas como lo son en las células bacterianas.
Desafortunadamente, los detalles de la terminación de la transcripción en eucariotas no se entienden tan bien como lo son en bacterias. Por lo tanto, nos centraremos en la iniciación, y luego consideraremos el procesamiento de diferentes ARN eucariotas en moléculas listas para usar.
1. Transcripción de ARNm eucariota
Los múltiples pasos de la transcripción de ARNm eucariota se muestran en la página siguiente.
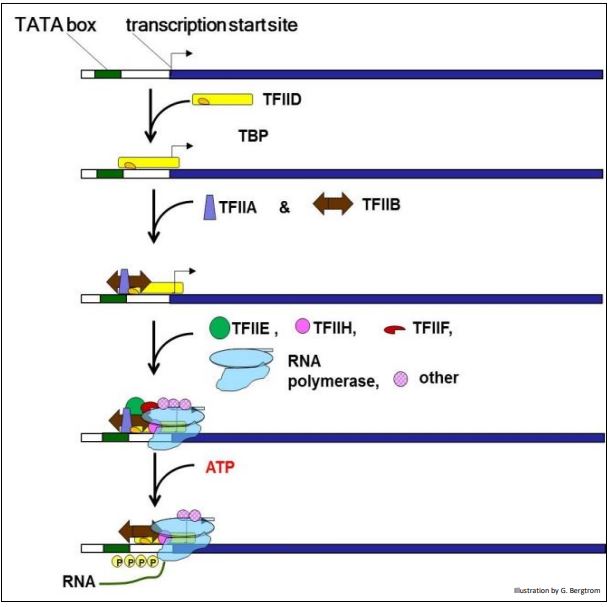
La transcripción de ARNm eucariotas por ARN polimerasa II comienza con el ensamblaje secuencial de un complejo de iniciación eucariota en un promotor génico. El promotor eucariota típico para un gen que codifica proteínas contiene un motivo de secuencia de ADN de caja TATA así como secuencias cadena arriba cortas adicionales. La proteína de unión a TATA (TBP) primero se une a la caja TATA junto con TFIID (factor de iniciación de la transcripción IID).
Este intermedio recluta a TFIIA y TFIIB. A continuación, TFGIIE, TFIIF y TFIIH, varios otros factores de iniciación y la ARN polimerasa II se unen para formar el complejo de iniciación de la transcripción. La fosforilación agrega varios fosfatos al extremo amino de la ARN polimerasa, después de lo cual algunos de los TF se disocian del complejo de iniciación. El complejo ARN polimerasa-TF restante ahora puede comenzar a producir el ARNm.
A diferencia de la ARN polimerasa procariota, la ARN polimerasa II eucariota no tiene una actividad helicasa inherente. Para ello, la transcripción de genes eucariotas se basa en la proteína TFIIH de múltiples subunidades, en la que dos subunidades tienen actividad helicasa. Consistente con la relación más estrecha de las arqueas con los eucariotas (más bien con los procariotas), la iniciación de la transcripción del ARNm arqueal se asemeja a la de los eucariotas, aunque requiere menos factores de iniciación durante la formación de un complejo de iniciación.
192 Transcripción de ARNm eucariota
Una diferencia significativa entre la transcripción procariota y eucariota es que la ARN polimerasa y otras proteínas involucradas en un promotor génico no ven ADN desnudo. En cambio, deben reconocer secuencias específicas de ADN a través de proteínas de cromatina. Por otro lado, todas las proteínas que interactúan con el ADN tienen en común la necesidad de reconocer las secuencias de ADN a las que deben unirse..., dentro de la doble hélice. Es decir, deben ver las bases dentro de la hélice, y no en su superficie de estructura de fosfato uniformemente electronegativa. Para ello, deben penetrar el ADN, generalmente a través del surco mayor de la doble hélice. ¡Veremos que las proteínas reguladoras del ADN enfrentan los mismos problemas para lograr interacciones específicas basadas en la forma!
193 Reconocimiento de Factores de Transcripción en Promotores
2. Transcripción eucariota de ARNt y 5SRNA
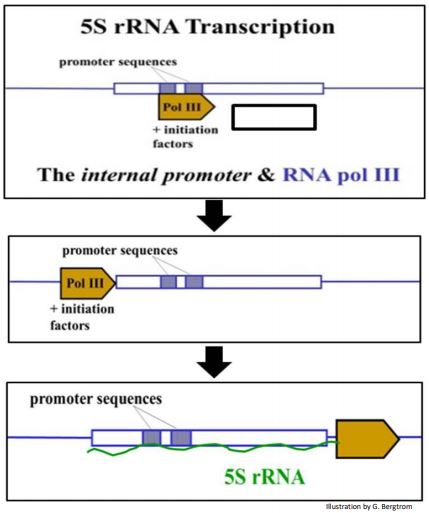
La transcripción de ARNr 5S y ARNt por la ARN polimerasa III es inusual ya que la secuencia promotora a la que se une (con la ayuda de factores de iniciación) no está aguas arriba de la secuencia transcrita, sino que se encuentra dentro de la secuencia transcrita. Después de unirse a este promotor interno, la polimerasa se reposiciona para transcribir el ARN desde el sitio de inicio de la transcripción para que el transcrito final contenga así la secuencia promotora.
A continuación se muestra el ARNr 5S por ARN polimerasa III.
3. Transcripción de los otros ARNr eucariotas
Los ARNt también son transcritos por la ARN polimerasa III de la misma manera que el ARNr 5S. Los otros ARNr son transcritos por la ARN polimerasa I, que se une a un promotor aguas arriba junto con factores de iniciación de la transcripción. Conocemos menos los detalles de este proceso en comparación con nuestra comprensión de la transcripción de ARNm. Exploraremos lo que sabemos a continuación. Como ya se señaló, la terminación de la transcripción no se entiende tan bien en eucariotas como en procariotas. Las etapas de terminación acoplada y poliadenilación comunes a la mayoría de los ARNm procariotas se discuten con más detalle a continuación, con un resumen útil en el sitio web de NIH-NCBI aquí.