
12.4: Regulación génica en eucariotas
- Page ID
- 54470

A. La diferencia entre la regulación génica eucariota y procariota
Recordemos un experimento descrito anteriormente e ilustrado a continuación.
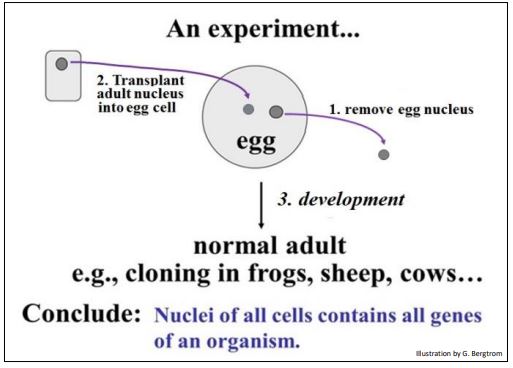
Los resultados de este experimento proporcionaron la evidencia de que incluso células muy diferentes de un organismo contienen los mismos genes. De hecho, en cualquier organismo eucariota multicelular, cada célula contiene el mismo ADN (genes). Por lo tanto, los diferentes tipos de células en un organismo no difieren en qué genes contienen, ¡sino en qué conjuntos de genes expresan! Mirado de otra manera, las células se diferencian cuando encienden nuevos genes y apagan los viejos. Así, la regulación génica produce diferentes conjuntos de productos génicos durante la diferenciación, lo que lleva a células que se ven y funcionan de manera diferente en el organismo.
220 Un experimento: todas las células de un organismo tienen el mismo genoma
En comparación con los procariotas, muchos pasos en eucariotas se encuentran entre la transcripción de un ARNm y la acumulación de un producto final polipeptídico. Once de estos pasos se muestran en la ruta de gen a proteína a continuación.
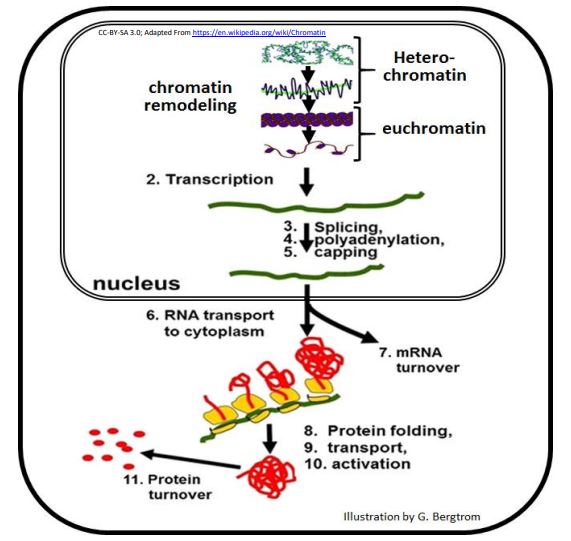
Teóricamente, las células podrían encenderse, apagarse, acelerar o ralentizar cualquiera de los pasos de esta vía, cambiando la concentración en estado estacionario de un polipéptido en las células. Si bien la regulación de cualquiera de estas etapas es posible, la expresión de un solo gen se controla típicamente en solo uno o unos pocos pasos. Una forma común de regulación génica es a nivel de iniciación de la transcripción, similar al control transcripcional en bacterias, en principio si no en detalle.
B. Complejidades de la Regulación Génica Eucariota
La regulación génica en eucariotas es más compleja que en procariotas. Esto se debe en parte a que sus genomas son más grandes y porque codifican más genes. Por ejemplo, el genoma de E. coli alberga alrededor de 5,000 genes, en comparación con alrededor de 25 mil genes en humanos. Además, los eucariotas pueden producir incluso más de 25,000 proteínas mediante el corte y empalme alternativo de ARNm y en al menos algunos casos, iniciando la transcripción desde sitios de inicio alternativos en el mismo gen. Y por supuesto, ¡la actividad de muchos más genes debe coordinarse sin el beneficio de los operones multigénicos! Finalmente, la regulación de genes eucariotas se complica porque todo el ADN nuclear está envuelto en proteína en forma de cromatina.
Todos los organismos controlan la actividad génica con factores de transcripción que se unen a secuencias específicas de ADN (elementos reguladores cis). En eucariotas, estos elementos pueden ser proximales a (cerca) del promotor de un gen, o distales a (bastante lejos de) el gen que regulan. A continuación se muestra un mapa eucariota que muestra los componentes de un gen típico y sus elementos reguladores asociados que actúan en cis.
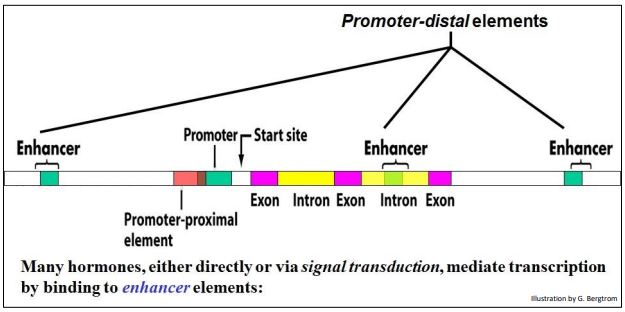
Los potenciadores son elementos cis distales típicos que reconocen y se unen a factores de transcripción para aumentar la tasa de transcripción de un gen. Por extraño que parezca, estos elementos cortos de ADN pueden estar en la región 5' o 3' no traducida del gen, o incluso dentro de intrones, y pueden estar a miles de pares de bases lejos de los genes que controlan. Tenga en cuenta que los elementos potenciadores están incluso en intrones también pueden estar muy lejos del sitio de inicio de la transcripción de un gen.
Las regiones reguladoras aguas arriba de genes eucariotas (a la izquierda de un promotor génico como se muestra anteriormente) a menudo tienen sitios de unión distales para más de unos pocos factores de transcripción, algunos con efectos positivos (potenciadores) y otros con efectos negativos (silenciamiento). Por supuesto, cuáles de estas regiones de ADN son activas en el control de un gen depende de qué factor o factores de transcripción estén presentes en el núcleo. Conjuntos de reguladores positivos trabajarán juntos para coordinar y maximizar la expresión génica cuando sea necesario, y conjuntos de reguladores negativos unirán elementos reguladores negativos para silenciar un gen.
222 Los factores de transcripción se unen al ADN cerca y lejos
Vimos que en eucariotas, el inicio de la transcripción involucra muchos factores de transcripción y la ARN polimerasa II actuando en un promotor génico para formar un complejo de preiniciación de la transcripción. La proteína de unión a TFIID o TATA es uno de los primeros factores en doblarse, haciendo que el ADN en la región promotora se doble, al igual que la proteína CAP en las bacterias. TFIID también recluta otros factores de transcripción para el promotor. Al igual que en las bacterias, doblar el ADN afloja los enlaces H entre las bases, facilitando el desenrollamiento de la doble hélice cerca del gen. Doblar el ADN eucariota también trae proteínas reguladoras distales unidas a secuencias potenciadoras lejos del promotor junto con las proteínas unidas a elementos reguladores más proximales, como se muestra en el dibujo a continuación.
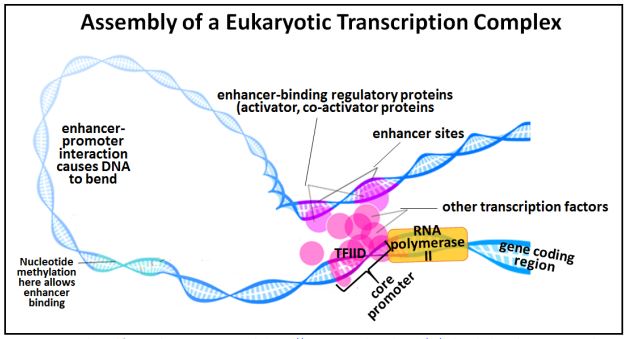
Los sitios de metiación de nucleótidos pueden facilitar la unión reguladora de proteína-potenciador. Cuando tales proteínas reguladoras, aquí llamadas activadoras (es decir, de transcripción), se unen a sus potenciadores, adquieren una afinidad por cofactores proteicos que permiten el reconocimiento y la unión a otras proteínas en el complejo de iniciación de la transcripción. Esta atracción estabiliza la curva en el ADN que luego facilita que la ARN polimerasa II inicie la transcripción
223 Ensamblaje de un complejo eucariota de iniciación de transcripción
Vale la pena recordarnos que es la forma y el cambio alostérico los que permiten las interacciones de DNaproteína (de hecho, cualquier interacción de macromoléculas). El represor lac que vimos anteriormente es un factor de transcripción con motivos de unión a ADN de hélice-giro-hélice. Este motivo y otros dos (dedo de zinc y cremallera de leucina) caracterizan a las proteínas de unión al ADN se ilustran a continuación.
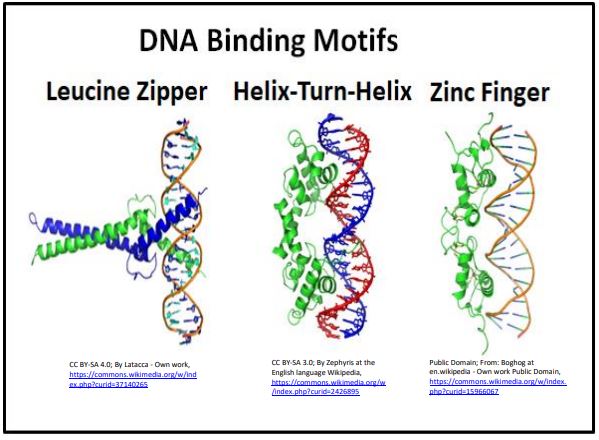
Los motivos de unión al ADN en cada proteína reguladora mostrada aquí se unen a uno o más elementos reguladores “visibles” al factor de transcripción en el surco mayor de la doble hélice.
224 Dominios/motivos del factor de transcripción se unen a secuencias de ADN específicas
A continuación veremos algunas formas comunes en las que las células eucariotas son señaladas para activar o desactivar los genes, o para aumentar o disminuir sus tasas de transcripción. Como describimos estos modelos, recuerde que las células eucariotas regulan la expresión génica en respuesta a cambios en ambientes extracelulares. Estos pueden ser cambios impredecibles, no programados en la composición de la sangre o del líquido extracelular (iones, pequeños metabolitos), o dictados por cambios en un programa genético de diferenciación y desarrollo a largo plazo. Los cambios en la expresión génica incluso obedecen a los ritmos circadianos (diarios), el tictac de un reloj. En los eucariotas, los cambios en la expresión génica, esperados o no, suelen estar mediados por la liberación oportuna de señales químicas de células especializadas (e.g., hormonas, citocinas, factores de crecimiento, etc.). Nos centraremos en algunos modelos mejor entendidos de regulación génica por estas señales químicas.
C. Regulación de la expresión génica por las hormonas que ingresan a las células y las que no
Los elementos reguladores de genes (cis) en el ADN y los factores de transcripción que se unen a ellos han coevolucionado. ¡Pero no sólo eso! Los organismos eucariotas han desarrollado vías completas que responden a señales de desarrollo ambientales o programadas y conducen a una respuesta celular apropiada. Los químicos que regulan los genes en procariotas no suelen ser señales comunicadas por otras células. En los eucariotas, los químicos liberados por algunas células señalan a otras células para que respondan, coordinando así la actividad de todo el organismo. Las hormonas liberadas por las células en las glándulas endocrinas son moléculas de señal bien conocidas; las hormonas afectan a las células diana en otras partes del cuerpo
225 Químicos Que Controlan La Expresión Génica
1. Cómo las hormonas esteroides regulan la transcripción
Las hormonas esteroides cruzan las membranas celulares para tener sus efectos. Las hormonas esteroides comunes incluyen testosterona, estrógenos, progesterona, glucocorticoides y corticoides minerales. Una vez en la célula diana, tales hormonas se unen a una proteína receptora de hormonas esteroideas para formar un complejo hormona-receptor esteroideo. El receptor puede estar en el citoplasma o en el núcleo, pero al final, el complejo hormona-receptor debe unirse a elementos reguladores de ADN de un gen para potenciar o silenciar la transcripción. Por lo tanto, una hormona esteroide debe cruzar la membrana plasmática, y también puede necesitar cruzar la envoltura nuclear.
Siga el enlace de una hormona esteroide a un receptor citoplásmico a continuación.
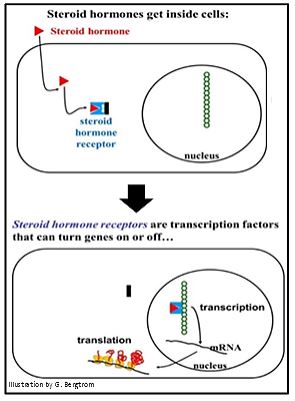
Aquí la hormona (el triángulo) entra en la célula. Un cambio alostérico en el receptor libera una subunidad proteica llamada Hsp90 (el rectángulo negro en la ilustración). El receptor unido a hormonas restante ingresa al núcleo.
Lo fascinante de Hsp90 es que se descubrió por primera vez en celdas sometidas a estrés térmico. Cuando la temperatura se eleva lo suficiente, las células cierran la mayor parte de la transcripción y en su lugar transcriben Hsp 90 y/u otros genes especiales de choque térmico. Las proteínas resultantes del choque térmico parecen proteger a las células contra el daño metabólico hasta que las temperaturas vuelven a la normalidad. Dado que la mayoría de las células nunca experimentan temperaturas tan altas, la importancia evolutiva de este mecanismo protector no está clara. Como ahora sabemos, las proteínas de choque térmico tienen funciones celulares críticas, en este caso bloqueando el sitio de unión al ADN de un receptor hormonal hasta que una hormona esteroide específica se una a él.
¡Regresa a la acción hormonal! Ya no asociado con la proteína Hsp90, el receptor unido a su cofactor hormonal se une a un elemento de control de la transcripción que actúa en cis en el ADN, activando o desactivando la transcripción de un gen. Los receptores hormonales para algunas hormonas esteroides ya están en el núcleo de la célula, por lo que la hormona debe cruzar no sólo la membrana plasmática, sino también la envoltura nuclear para acceder al receptor.
En cuanto a las funciones de las hormonas esteroides, ya vimos que los glucocorticoides encienden los genes de la gluconeogénesis. Las hormonas esteroides también controlan el desarrollo sexual y el ciclo reproductivo en las hembras, la homeostasis de sal y mineral en la sangre, la metamorfosis en artrópodos, etc., todo regulando la expresión génica.
2. Cómo las hormonas proteicas regulan la transcripción
Las hormonas proteicas son, por supuesto, grandes y solubles, con superficies altamente cargadas. Otras hormonas pueden ser relativamente pequeñas (p. ej., adrenalina), pero están cargadas. Las moléculas de señal grandes o altamente cargadas no pueden atravesar la barrera de fosfolípidos de la membrana plasmática. Para tener algún efecto en absoluto, deben unirse a receptores en la superficie de las células. Estos receptores son típicamente glicoproteínas de membrana.
La información (señales) transportadas por las hormonas proteicas deben ser transportadas a la célula indirectamente, mediante un proceso llamado transducción de señales. Existen dos vías bien conocidas de transducción de señales, cada una de las cuales implica activar vías de fosforilación de proteínas en el citoplasma. La cascada de fosforilación que resulta activa un factor de transcripción que se une al ADN regulador, activando o apagando un gen.
La unión de una hormona a un receptor de superficie celular conduce a un cambio alostérico en el receptor. Esto a su vez activa otras proteínas ya sea en la membrana plasmática o en el citoplasma, conduciendo a la síntesis de un segundo mensajero citoplásmico. El segundo mensajero típicamente se une a una proteína quinasa en el citoplasma, lanzando una serie de fosforilaciones de proteínas, o una cascada de fosforilación. El último de la serie de proteínas a fosforilar es un factor de transcripción activado que se unirá a una secuencia de ADN cis-reguladora.
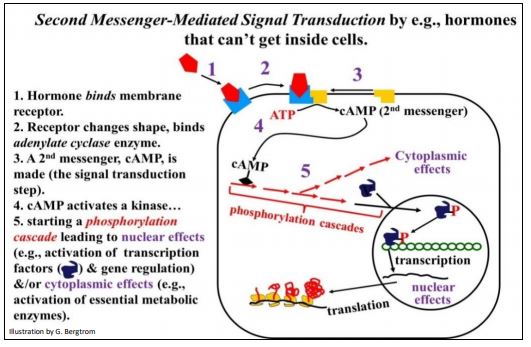
El AMPc fue el primer metabolito segundo mensajero en ser descubierto. Intermedia en muchas respuestas hormonales, controlando tanto la actividad génica como la actividad enzimática. El cAMP se forma cuando el receptor hormonal en la membrana se une y activa una enzima adenilato ciclasa unida a la membrana. El AMPc producido luego se une a una proteína quinasa, la primera de varias en una cascada de fosforilación. La transducción de señales mediada por AMPc se resume en la siguiente ilustración.
Un tipo diferente de transducción de señales implica un receptor hormonal que es en sí mismo la proteína quinasa. El papel de los receptores hormonales ligados a enzimas en la transducción de señales se resume a continuación.
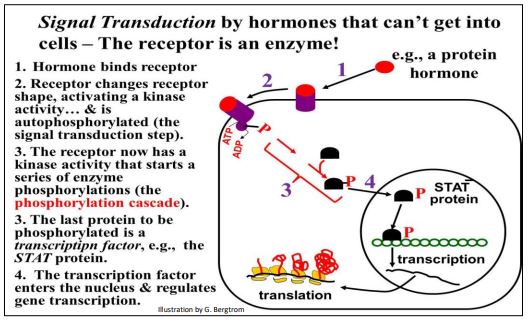
La unión de la proteína señal (por ejemplo, hormona) al receptor ligado a enzimas provoca un cambio alostérico que activa el receptor quinasa, iniciando la cascada de fosforilación dando como resultado un factor de transcripción activo. Analizamos la transducción de señales con más detalle en otro capítulo.
227 La transducción de señales puede conducir a la regulación génica
D. Regulación de genes eucariotas significa contender con la cromatina
Consideremos nuevamente la ilustración de los diferentes niveles de estructura de cromatina (abajo).
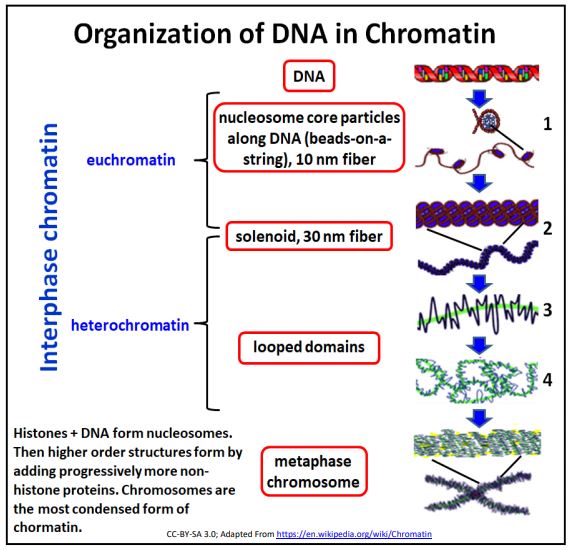
Los factores de transcripción se unen a secuencias de ADN específicas detectándolas a través de los surcos (principalmente el surco mayor) en la doble hélice. El dibujo anterior nos recuerda sin embargo, que a diferencia del ADN casi desnudo de las bacterias, el ADN eucariota (nuclear) está recubierto con proteínas que, en conjunto son en masa, mayores que la masa de ADN que cubren. El complejo proteína-ADN del genoma es, por supuesto, la cromatina.
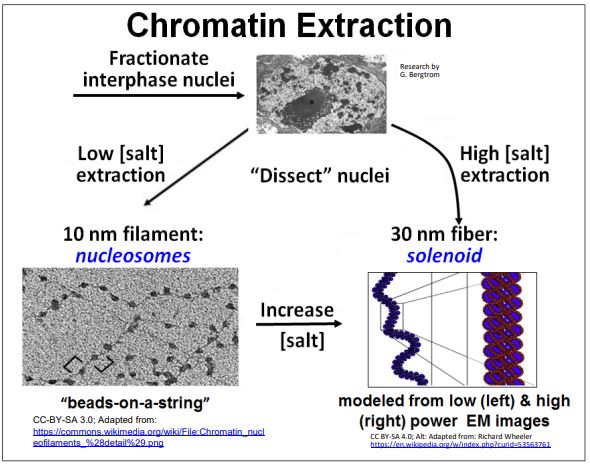
Nuevamente, como recordatorio, el ADN recubierto con proteínas histonas forma la estructura de perlas sobre una cadena de 9 nm de diámetro en la que las perlas son los nucleosomas. La asociación de proteínas específicas no histonas hace que los nucleosomas se plieguen sobre sí mismos para formar el solenoide de 30 nm. Como vimos antes, es posible extraer selectivamente la cromatina. Dé un segundo vistazo a los resultados de extracciones típicas de cromatina de núcleos aislados a continuación.
La acumulación adicional de proteínas no histonas conduce a un mayor plegamiento y a la formación de eucromatina y heterocromatina características de las células que no se dividen. En las células en división, la cromatina se condensa aún más para formar los cromosomas que se separan durante la mitosis o la meiosis.
Recordemos que el análisis bioquímico del extracto de filamento de 10 nm reveló que el ADN se envuelve alrededor de octámeros de proteínas histonas, los nucleosomas o perlas en esta estructura de perlas sobre astring. Las proteínas histonas están altamente conservadas en la evolución eucariota (no se encuentran en procariotas). También son muy básicos (muchos residuos de lisina y arginina) y por lo tanto muy positivamente cargados. Esto explica por qué son capaces de organizarse uniformemente a lo largo del ADN, uniéndose al esqueleto fosfodiéster cargado negativamente del ADN en la doble hélice.
Dado que el ADN en la eucromatina está menos apretado que en la heterocromatina, quizás los genes activos se encuentran en la eucromatina y no en la heterocromatina. Los experimentos en los que se aislaron extractos de cromatina nuclear total y se trataron con la enzima desoxirribonucleasa (DNasa) revelaron que el ADN en los genes activos se degradó más rápidamente que el ADN no transcrito. Más detalles sobre estos experimentos se pueden encontrar en los dos enlaces a continuación.
228 Pregunta: ¿Se transcribe el ADN eucromático?
229 Experimento y Respuesta: La eucromatina se transcribe
Los resultados de tales experimentos son consistentes con la sugerencia de que los genes activos son más accesibles a la DNasa porque están en cromatina menos enrollada o menos condensada. El ADN en la cromatina más condensada está rodeado por más proteínas y, por lo tanto, es menos accesible y protegido del ataque de DNasa. Cuando se empaquetan en cromosomas durante la mitosis o meiosis, todos los genes están en gran parte inactivos.
La regulación de la transcripción génica debe ocurrir en células que no se dividen o durante la interfase de las células, donde es posible cambiar la forma de la cromatina (remodelación de la cromatina) para silenciar algunos y activar otros genes. Cambiar la conformación de la cromatina implica la modificación química de proteínas y ADN de la cromatina. Por ejemplo, la cromatina puede modificarse por acetilación de histonas, desacetilación, metilación y fosforilación, reacciones catalizadas por histona acetiltransferasas (enzimas HAT), desacetilasas, metiltransferasas y quinasas, respectivamente. Por ejemplo, la acetilación de lisinas cerca del extremo amino de las histonas H2B y H4 tiende a desenrollar nucleosomas y abrir el ADN subyacente para su transcripción. La desacetilación entonces, promueve la condensación de la cromatina en las regiones afectadas del ADN. Así mismo, la metilación de lisinas o argininas (¡los aminoácidos básicos que caracterizan a las histonas!) de H3 y H4 pueden abrir ADN para la transcripción, mientras que la desmetilación tiene el efecto contrario. En un caso, la di-metilación de una lisina en H3 puede suprimir la transcripción. Estas modificaciones químicas afectan el reclutamiento de otras proteínas que alteran la conformación de la cromatina y finalmente activan o bloquean la transcripción.
Este reversible y su efecto sobre la cromatina se ilustran a continuación.
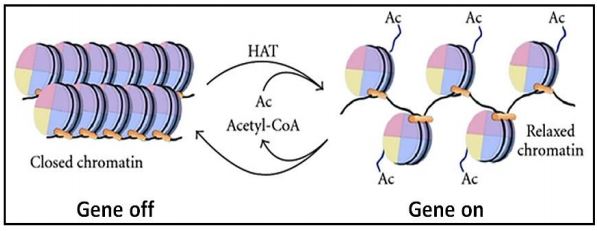
Los nucleosomas en sí mismos pueden ser movidos, deslizados y reposicionados de otra manera por complejos que hidrolizan ATP para obtener energía para lograr los cambios físicos. Algunos cánceres están asociados con mutaciones en genes para proteínas involucradas en la remodelación de la cromatina. Esto sin duda, ya que las fallas de la remodelación normal podrían afectar negativamente el ciclo celular normal y la replicación normal. De hecho, un único patrón específico de metilación puede marcar el ADN en múltiples tipos de cáncer (consulte Cinco cánceres con la misma firma genómica - Implicaciones).
E. Regulación de todos los genes en un cromosoma a la vez
Recordemos que los cromosomas X en las células somáticas femeninas humanas están inactivados, visibles en el núcleo como un cuerpo Barr. Uno de los dos cromosomas X en las moscas femeninas de la fruta también es inactiva. Sin embargo, tanto machos como hembras de Drosophila (¡presumiblemente también nosotros!) requieren la expresión del gen del cromosoma X durante la embriogénesis. Dada la diferencia en la dosis del gen del cromosoma X entre hombres y mujeres, ¿los machos se las arreglan con menos productos génicos del cromosoma X que las mujeres?
Los experimentos que observaron la expresión del gen del cromosoma X en moscas masculinas y femeninas revelaron niveles similares de productos génicos. Resulta que la actividad de un cuerpo nuclear llamado HLB (H istone L ocus B ody) es necesaria para aumentar la transcripción del gen del cromosoma X. Se demostró que una proteína llamada CLAMP (C hromatin- L entintado A daptor para M ale-specific letal (MSL) P roteína), se une a repeticiones de nucleótidos GAGA que se encuentran entre los genes de las histonas 3 y 4. Como hay alrededor de 100 repeticiones del locus de histonas de cinco genes en los cromosomas X, y por lo tanto alrededor de 100 repeticiones de las repeticiones GAGA. Por lo tanto, muchas proteínas CLAMP se unen a los HLB, donde reclutan muchas proteínas MSL. Los complejos proteicos MSL que se forman entonces aumentan globalmente la expresión del gen del cromosoma X masculino, compensando la menor dosis del gen X en los machos. Lea la investigación original aquí (L.E. Reider et al. (2018) Genes & Development 31:1-15). Y finalmente, ¡hay evidencia emergente de que la acción HLB también puede estar involucrada en la inactivación de todo un cromosoma X femenino más tarde en la embriogénesis en las hembras!