
17.7:17.7 Transducción de Señal
- Page ID
- 54443

Cuando las moléculas efectoras químicas hidrofóbicas, como las hormonas esteroides, alcanzan una célula diana, pueden cruzar la membrana hidrófoba y unirse a un receptor intracelular para iniciar una respuesta. Cuando moléculas efectoras grandes (por ejemplo, hormonas proteicas) o hormonas altamente polares (p. ej., adrenalina) alcanzan una célula diana, no pueden cruzar la membrana celular. En cambio, se unen a receptores de proteínas transmembrana en las superficies celulares. Un cambio conformacional iniciado en el dominio extracelular del receptor induce un cambio alostérico adicional en el dominio citoplásmico del receptor. Una serie secuencial de eventos moleculares convierte entonces la información entregada por el efector externo en información intracelular, un proceso llamado transducción de señales. A continuación se ilustra un esquema general de los eventos de transducción de señales.
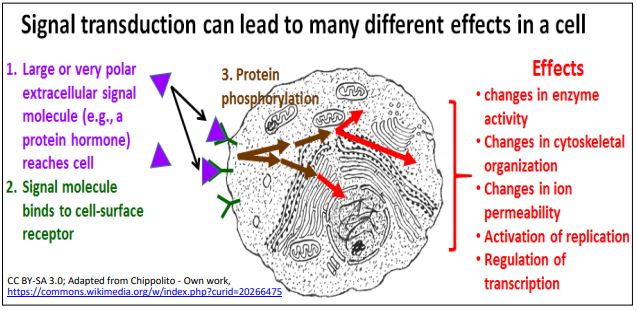
Muchos efectos de la transducción de señales están mediados por una secuencia, o cascada de fosforilaciones de proteínas catalizadas por proteínas quinasas dentro de la célula. Aquí consideraremos los receptores ligados a proteína G y ligados a enzimas.
315 Introducción a la Transducción de Señales
A. Transducción de señales mediada por proteína G por PKA (proteína quinasa A)
Las proteínas de unión a GTP (G-Proteínas) transducen señales extracelulares induciendo la producción de moléculas de segundo mensajero en las células. Cuando las hormonas u otras moléculas efectoras (señal) se unen a sus receptores de membrana, un cambio alostérico en el dominio citoplásmico del receptor aumenta la afinidad del dominio citoplásmico el receptor por las proteínas G en la superficie interna de la membrana plasmática. Las proteínas G son trímeros que consisten en\(\alpha \),\(\beta \) y\(\gamma \) subunidades, incrustados en la superficie citoplásmica de las membranas celulares sensibles. La transducción de señales mediada por proteína G se ilustra en los siete pasos que se muestran en la página siguiente.
El receptor cambia de forma al unirse a su molécula señal efectora (etapas 1, 2). En esta conformación, el receptor reconoce y se une al trímero de proteína G en la superficie citoplásmica de la membrana plasmática (paso 3). Al unirse el trímero al receptor, el GTP desplaza el GDP sobre la \(\alpha \)subunidad de la proteína G (paso 4).
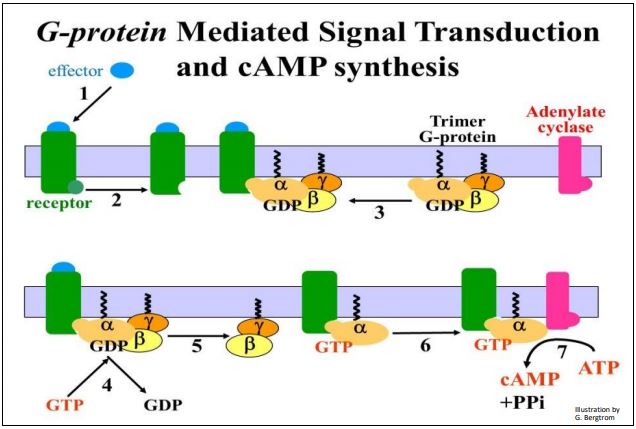
Después de un cambio conformacional, la \(\alpha \)subunidad se disocia de las\(\gamma \) subunidades\(\beta \) y (paso 5). En esta ilustración, la\(\alpha \) subunidad GTP- ahora puede unirse a una enzima transmembrana, la adenilato ciclasa (etapa 6). Finalmente, la señal química extracelular inicial se transduce a una respuesta intracelular que involucra moléculas de segundo mensajero (etapa 7). En este caso, el segundo mensajero es cAMP. La conocida respuesta de lucha o huida a la adrenalina en las células hepáticas de animales superiores es un buen ejemplo de una respuesta celular mediada por cAMP. Después de que la adrenalina se une a sus receptores, las proteínas G a su vez se unen al lado citoplásmico del receptor, que luego se une a la adenilato ciclasa. El cAMP se une y activa la proteína quinasa A (PKA), desencadenando la respuesta en cascada de amplificación. Algunos detalles de una cascada de amplificación de señal mediada por proteína G se detallan en la ilustración de la página siguiente.
Después de la activación de la adenilato ciclasa (etapas 1 y 2 en el dibujo), se sintetiza AMPc y se une a dos de las cuatro subunidades de una PKA inactiva (etapa 3). Un cambio conformacional disocia el tetrámero en dos subunidades inertes unidas a AMPc y dos subunidades PKA activas (paso 4). Cada enzima PKA activa cataliza la fosforilación y activación de una enzima llamada fosforilasa quinasa (paso 5).
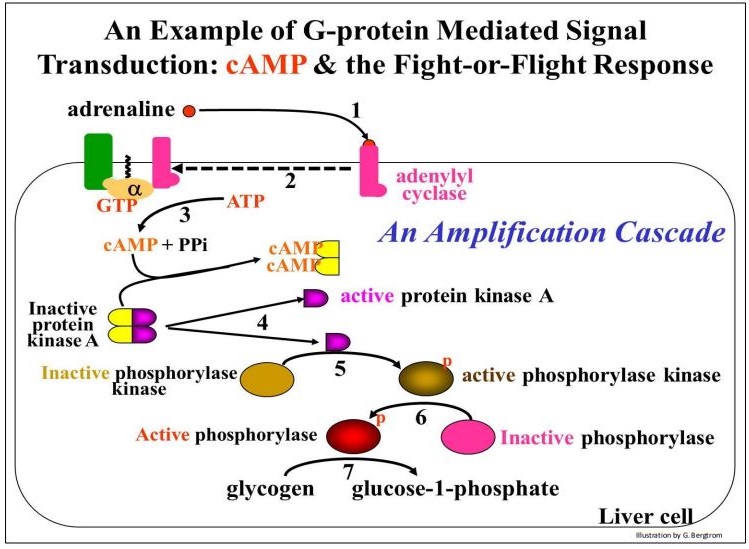
En la etapa 6, la fosforilasa quinasa cataliza la fosforilación de glucógeno fosforilasa. Finalmente, al final de la cascada de fosforilación, la glucógeno fosforilasa ahora activa cataliza la hidrólisis del glucógeno a glucosa-1-fosfato (paso 7). Esto da como resultado una rápida recuperación de glucosa libre de las células hepáticas a la circulación. Recuerda cómo funciona esto revisando la conversión de glucosa-1 fosfato (G-1-P) a G-6-P en la glucólisis y su destino en la gluconeogénesis. Por supuesto, el aumento en la glucosa circulante proporciona la energía para la decisión de luchar o huir.
317 Activación de Proteína G de la Proteína Quinasa A y una Respuesta de Lucha o Huida
Además de activar enzimas que descomponen el glucógeno, la PKA activada por AMPc media las respuestas celulares a diferentes efectores dando como resultado una cascada de fosforilación que conduce a
- Activación de enzimas que catalizan la síntesis de glucógeno.
- Activación de lipasas que hidrolizan ácidos grasos a partir de triglicéridos.
- Montaje de microtúbulos.
- Desmontaje de microtúbulos.
- Efectos mitogénicos (activación de enzimas de replicación).
- Activación de factores de transcripción aumento/decreciente de la expresión génica.
Por supuesto, cuando la respuesta celular ya no es necesaria por el organismo, debe dejar de producir las moléculas señal (hormona u otro efector). A medida que bajan sus niveles, las moléculas efectoras se disocian de sus receptores y la respuesta se detiene. ¡Todo esto es posible porque la unión de señales a sus receptores es libremente reversible! Esto está animado para la transducción de señales basada en proteínas G en el siguiente enlace.
B. Transducción de señales mediante PKC
Muchas respuestas que involucran proteínas G comienzan activando la adenilato ciclasa de membrana integral. Una vía de señalización mediada por proteína G diferente genera otros segundos mensajeros. La proteína quinasa C (PKC) juega un papel importante en la activación de estos otros segundos mensajeros y posteriores cascadas de fosforilación en las que la activación de solo unas pocas moléculas enzimáticas en la célula da como resultado la activación de muchas más enzimas. Al igual que PKA, la transducción de señales mediada por PKC también amplifica la primera respuesta molecular de la célula al efector. El papel de las proteínas G es similar para la transducción de señales PKA y PKC. Las respuestas pueden incluir diversos efectos en diferentes células..., o incluso en las mismas células usando diferentes señales efectoras. Pero, la transducción de señales PKC y PKA difieren en que la activación de PKC requiere un paso adicional, así como la generación de dos moléculas mensajeras intracelulares. Los eventos que condujeron a la activación de PKC se ilustran a continuación.
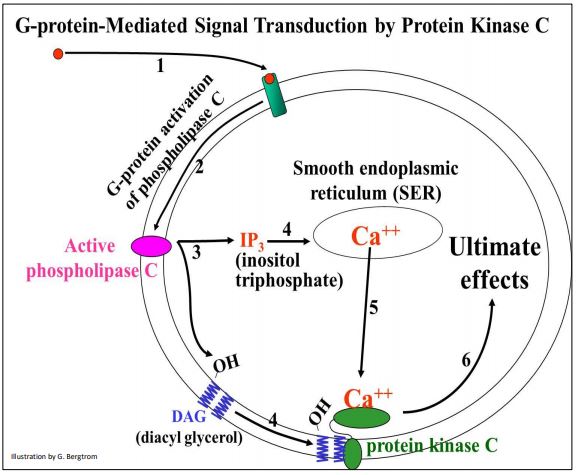
Aquí están los detalles de los pasos que conducen a la activación de PKC. Una molécula señal efectora se une a su receptor, activando una enzima fosfolipasa C de membrana integral. La fosfolipasa C cataliza la formación de inositol trifosfato citosólico (IP 3) y diacil glicerol unido a membrana (DAG), dos de esas otras moléculas intracelulares de segundo mensajero. IP3 interactúa con receptores en el retículo endoplásmico liso, provocando la liberación de iones Ca++ secuestrados en el citoplasma. Finalmente, los iones Ca 2+ y DAG activan la Proteína Quinasa C (PKC) que luego inicia una cascada de amplificación de fosforilación liderando respuestas específicas de células.
318 Activación de Proteína G de Proteína Cinasa C y Fosfolipasa C
Los efectos mediados por la proteína quinasa C incluyen:
- Liberación de neurotransmisores.
- Hormona (hormona del crecimiento, hormona leutinizante, testosterona) secreción que conduce al crecimiento celular, división y diferenciación.
- Hidrólisis glucógeno, síntesis de grasas.
Los efectos adicionales de la fosfolipasa C independientes incluyen:
- Desglose del glicógeno hepático.
- Secreción de amilasa pancreática.
- Agregación plaquetaria.
PKA y PKC son serina-treonina quinasas que colocan fosfatos sobre serina o treonina en polipéptidos diana. Consideremos a continuación las tirosina quinasas.
C. Transducción de señales mediada por receptor tirosina quinasa
La actividad intracelular de estos receptores se encuentra en el dominio citoplasmático del propio receptor. Cuando se une a su efector, el receptor-quinasa cataliza la fosforilación de aminoácidos tirosina específicos en proteínas diana. Mientras estudiaban la acción del factor de crecimiento nervioso (NGF) y del factor de crecimiento epidérmico (EGF) en la estimulación del crecimiento y la diferenciación de nervios y piel, Stanley Cohen y Rita Levi-Montalcini descubrieron el receptor EGF, la primera enzima -ligada a la tirosina quinasa..., ¡y ganó el Premio Nobel de Fisiología o Medicina en 1986! Observe la animación de la transducción de señales del receptor quinasa en el siguiente enlace (se proporciona una descripción en los siguientes párrafos).
319 Transducción de señal de receptor quinasa
Las quinasas receptoras de membrana monomérica se dimerizan cuando se unen a ligandos efectores, momento en el que las proteínas SH 2 que contienen grupos sulfhidrilo se unen a cada monómero. Esto activa el dominio quinasa del receptor. Después de múltiples fosforilaciones cruzadas de los monómeros receptores, las proteínas SH 2 caen permitiendo que los receptores interactúen con otras proteínas citoplásmicas para continuar la vía de respuesta. La respuesta característica a la señalización de EGF y NGF es la proliferación celular. No es sorprendente que las mutaciones correlacionadas con las células cancerosas a menudo se encuentran en vías de señalización que conducen a la proliferación celular (crecimiento y división). Los genes causantes de cáncer, u oncogenes, en realidad se descubrieron por primera vez en los virus, pero J. Michael Bishop y Harold Varmus ganaron el Premio Nobel de Fisiología o Medicina en 1964 por demostrar que las células eran en realidad el origen de un retrovirus de pollo (el Virus del Sarcoma de Rous). Los oncogenes resultan ser mutaciones de genes para proteínas en vías de transducción de señales mitogénicas. En circunstancias normales, las señales químicas mitogénicas (como EGF) se unen a sus receptores e inducen a las células diana a comenzar a dividirse.
La activación mediada por la proteína Ras de una cascada de fosforilación que conduce a la MAP (proteína activada por mitógeno) quinasa es un ejemplo de tal vía de transducción de señales, una con un papel central en muchas vías de señalización de cinasas receptoras. El gen Ras fue uno de los originalmente descubiertos como un oncogén cuya mutación conduce a una división celular incontrolada, es decir, cáncer. ¡De hecho, la actividad del gen Ras/proteína puede ser responsable de hasta el 30% de todos los cánceres!
320 El Oncogén RAS, sus Efectos Mitogénicos Normales y Cáncer
La MAP quinasa fosforila factores de transcripción y otras proteínas nucleares que afectan la actividad génica, lo que lleva a la proliferación y diferenciación celular, como se muestra a continuación.
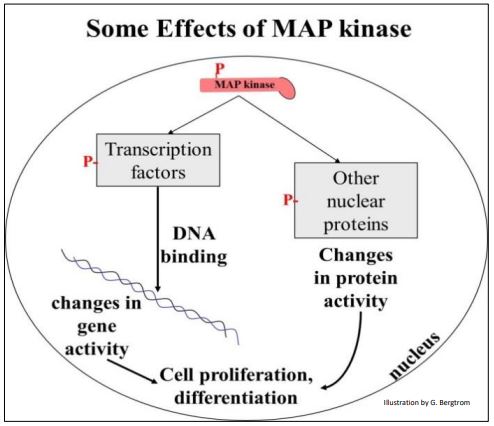
D. Transducción de señales en la evolución
Vimos que la transducción de señales normalmente toma algunas moléculas de señal que interactúan con unos pocos receptores de la superficie celular para amplificar una respuesta en una cascada de reacciones enzimáticas, típicamente fosforilaciones, para activar (o inactivar) proteínas diana. Las cascadas de amplificación pueden tomar una sola interacción efector-receptor y magnificar su efecto en la célula en órdenes de magnitud, haciendo que los sistemas de señalización sean rápidos y altamente eficientes. El rango de respuestas celulares y sistémicas (organizativas) a una misma señal química es amplio y complejo. Diferentes tipos de células pueden tener receptores para el mismo efector, pero responden de manera diferente. Por ejemplo, la adrenalina se dirige a células del hígado y vasos sanguíneos entre otras, con diferentes efectos en cada una. A medida que sucede, la adrenalina también es un neurotransmisor. Al parecer, a medida que los organismos evolucionaron, se volvieron más complejos en respuesta a los imperativos ambientales, adaptándose cooptando sistemas de señalización ya existentes al servicio de nuevas vías. Así como el mismo evento de transducción de señales puede conducir a diferentes vías de respuesta en diferentes células, la evolución ha permitido que diferentes vías de transducción de señales se involucren en diafonía. Esto es cuando dos vías de transducción de señales diferentes se cruzan en las mismas células. En un ejemplo, el AMPc producido en el extremo frontal de la vía de señalización de PKA puede activar (o en las circunstancias adecuadas, inhibir) enzimas en la ruta de MAP quinasa. Estos efectos dan como resultado cambios en los niveles de factores de transcripción activos o inactivos y, por lo tanto, pueden modular la expresión de un gen usando dos (o más) señales. Apenas estamos empezando a entender lo que parece menos un lineal.