
18.2: Componentes citoesqueléticos
- Page ID
- 53871

La mayoría de las células eucariotas parecen un saco de citoplasma unido a la membrana que contiene un núcleo y organelos variados en un microscopio óptico. A finales del siglo XIX, los microscopistas describieron una dramática reorganización estructural de las células en división. En la mitosis, los cromosomas duplicados (es decir, cromátidas) se condensan en el núcleo justo cuando la membrana nuclear se disuelve. Las fibras del huso emergen y luego parecen separar las cromátidas hacia polos opuestos de la celda. Las fibras del huso resultan ser haces de microtúbulos, cada uno de los cuales es un polímero de proteínas de tubulina. Veamos a continuación esa micrografía de fluorescencia de una célula de metafase mitosante nuevamente; la mayor parte de la célula que no sea la que está fluorescente no es visible en la micrografía.
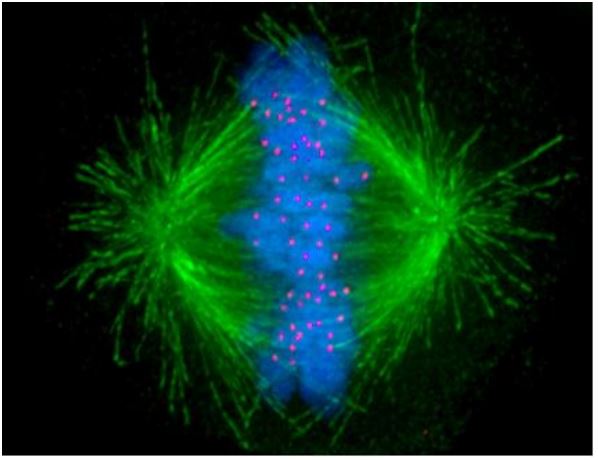
Para obtener esta imagen, se realizaron anticuerpos contra microtúbulos purificados, cinetocoros y proteínas cromosómicas (o ADN), y luego se unieron a diferentes fluoróforos (etiquetas fluorescentes moleculares orgánicas). Cuando los fluoróforos se agregaron a las células divisorias en metafase, se unieron a sus respectivas fibras. Tras la irradiación con luz UV, los fluoróforos emiten diferentes colores de luz visible, visibles en un microscopio de fluorescencia. Los microtúbulos son verdes, los cromosomas metafase son azules y los cinetocoros son rojos en la micrografía.
Tanto la mitosis como la meiosis son ejemplos muy visibles de movimientos dentro de las células, ambos ya descritos a finales del siglo XIX. En cuanto al movimiento en organismos enteros, los estudios de mediados del siglo XX se centraron en lo que las estrías (o franjas) que se ven en el músculo esquelético en el microscopio óptico podrían tener que ver con la contracción muscular. Las estrías resultaron estar compuestas por un complejo proteico originalmente llamado actomiosina (acto para activo; miosina para músculo). La microscopía electrónica reveló posteriormente que la actomiosina (o actinomiosina) está compuesta por filamentos delgados (actina) y filamentos gruesos (miosina) que se deslizan unos sobre otros durante la contracción muscular.
La microscopía electrónica también insinuó una estructura citoplásmica más compleja de las células en general. El citoesqueleto consiste en varillas finas y tubos en estados más o menos organizados que permean la célula. Los más abundantes de estos componentes citoesqueléticos son microfilamentos, microtúbulos y filamentos intermedios. Pero, incluso la miosina está presente en las células no musculares, aunque a concentraciones relativamente bajas. Los microtúbulos explican los movimientos cromosómicos de la mitosis y la meiosis, mientras que junto con los microfilamentos (es decir, actina), permiten el movimiento de orgánulos dentro de las células (es posible que hayas visto la transmisión citoplasmática de cloroplastos de Elodea en un ejercicio de laboratorio de biología). Los microtúbulos también subyacen a la motilidad basada en cilios y flagelos de células enteras como paramecio, ameba, fagocitos, etc., mientras que los microfilamentos de actina y la miosina permiten el movimiento muscular y, por lo tanto, ¡un mayor movimiento animal! Finalmente, el citoesqueleto es una estructura dinámica. Sus fibras no sólo dan cuenta de los movimientos de la división celular, sino que también dan a las células resistencia mecánica y formas únicas. Todas las fibras se pueden desmontar, volver a ensamblar y reorganizar, permitiendo que las células cambien de forma, por ejemplo, creando pseudopodos en células ameboides y fibras fusiformes de mitosis y meiosis. En este capítulo analizamos las bases moleculares de la estructura celular y las diferentes formas de motilidad celular.