
20.4: Orígenes de las moléculas orgánicas en una atmósfera no reductora
- Page ID
- 53928

Una atmósfera prebiótica no reductora se basa en varios supuestos: (1) La Tierra primitiva habría tenido una gravedad insuficiente para contener H 2 y otros gases ligeros; por lo tanto, la “desgasificación” habría resultado en una pérdida de H2 y otros agentes reductores de la atmósfera. (2) Evidencia geológica sugiere que los océanos y la corteza terrestre se formaron temprano en el Hadean Eon, solo unos cientos de millones de años después de la formación del planeta. (3) Los estudios de 4.4 mil millones de años (principios del Hadean Eon) cristales de circón australianos sugieren que su estado de oxidación es el mismo que las rocas modernas, es decir, que la atmósfera de los primeros Hadeanos era en gran parte N 2 y CO 2, ¡una claramente no reductora! A continuación se muestra una imagen en color de este circón australiano.

Entonces la vida pudo haber comenzado en un ambiente no reductor. Sin embargo, ¿hasta qué punto podemos fechar la aparición de las primeras células reales en la tierra? La evidencia geológica sólida de la vida real data de hace 3.5-3.95 mil millones de años (es decir, el Eón Arcaico). Existe evidencia más suave de vida microbiana en forma de grafito y otros restos 'posibles' tan antiguos como hace 4.1 mil millones de años, cerca del final del Eón Hadeano. Así, independientemente de si la vida comenzó hace 3.5 o incluso 4.1 mil millones de años, la evidencia sugiere que los inicios de la vida tuvieron que lidiar con un ambiente no reductor.
Antes de examinar más de cerca otras evidencias de orígenes de vida en condiciones no reductoras, consideremos la Panspermia, la posibilidad de que la vida viniera a la tierra de fuentes extraterrestres y una hipótesis relacionada de que las moléculas orgánicas prebióticas provenían de fuentes extraterrestres . Luego examinaremos cómo podrían haberse formado las células en entornos terrestres localizados y favorables.
A. Panspermia - un origen extraterrestre de la vida terrenal
Panspermia postula que la vida misma llegó a nuestro planeta en cometas o meteoritos. Dado que es poco probable que estos hayan sostenido la vida en el espacio, deben haber sido una especie de 'buzón' interestelar en el que se depositaron formas de vida inactivas. Las celdas de los buzones deben haber sido criptobióticas. Hoy en día existen ejemplos de criptobiosis (p. ej., esporas bacterianas, ¡camarones en salmuera!). Una vez entregados al ambiente amigable para la vida de la Tierra, tales organismos emergerían de la latencia, eventualmente poblando el planeta. Sin embargo, no hay evidencia de vida latente o criptobiótica en cometas o meteoritos, y ninguna evidencia sólida que apoye la Panspermia. Por otro lado, hay evidencias al menos consistentes con una fuente extraterrestre de moléculas orgánicas, y abundancia para soportar más orígenes terrestres de la vida. En cualquier caso, las nociones de Panspermia (e incluso fuentes extraterrestres de moléculas orgánicas) simplemente plantean la cuestión de las condiciones que habrían llevado al origen de la vida en otra parte.
Si bien la panspermia no es un escenario favorecido, sin embargo es intrigante, en el sentido de que está en línea la probabilidad de que las moléculas orgánicas se formaron poco después del Big Bang. Además, si las moléculas orgánicas preparadas y el agua estuvieran disponibles, podemos esperar (¡y muchos lo hacen!) que hay vida en otros planetas. Esta expectativa ha estimulado una seria discusión y financiamiento de programas que buscan señales de vida en otros planetas. Por ejemplo, la NASA financió la búsqueda (y descubrimiento de) señales de agua de Rover en Marte. Incluso apoyó la Búsqueda más terrestre de Inteligencia Extraterrestre (el programa SETI), basada en la suposición de que la vida no solo existe en otros lugares, sino que evolucionó habilidades de comunicación de alto nivel (¿y por qué no?)! Para una historia fascinante sobre meteoritos de Marte que contienen agua y que valen más que el oro, haga clic en Obsesión marciana.
B. Orígenes extraterrestres de moléculas orgánicas
Aunque la vida no nos llegara ya hecha, ¿podrían haber llegado moléculas orgánicas a la tierra desde el espacio exterior? Son abundantes, por ejemplo en nubes interestelares, y podrían haberse convertido en parte de la tierra a medida que el planeta se formó hace alrededor de 4.8 mil millones de años. Esto sugiere que no hubo necesidad de crearlos de novo. Una hipótesis sugiere que meteoritos, cometas y asteroides, conocidos por contener moléculas orgánicas, los trajeron aquí durante los impactos ardientes en nuestro planeta. Los bombardeos de cometas y meteoritos habrían sido comunes hace 3.8 o más mil millones de años. En este escenario la cuestión de cómo (¡no en la tierra! ) ¡la energía libre y los precursores moleculares inorgánicos reaccionaron para formar moléculas orgánicas es discutible!
Una hipótesis relacionada sugiere que esos impactos ardientes en sí mismos proporcionaron la energía libre necesaria para sintetizar las moléculas orgánicas a partir de las inorgánicas, un escenario de síntesis a la llegada. Con esta hipótesis, por un lado, volvemos a una sopa primordial oceánica orgánica. Por otro, algunos han sugerido que las moléculas orgánicas producidas de esta manera (sin mencionar ninguna forma de vida primordial) probablemente habrían sido destruidas por los mismos impactos continuos de cuerpos extraterrestres; presenciar la extinción relativamente reciente de dinosaurios por un impacto de asteroide frente a la costa de México unos 65. Hace 5 millones de años.
C. Orígenes moleculares orgánicos más cerca del hogar
En lo profundo de los océanos, lejos de los bombardeos meteóricos y de la energía libre desenfrenada de un cielo libre de oxígeno y sin ozono, los respiraderos hidrotermales de aguas profundas habrían estado arrojando moléculas reductoras (por ejemplo, H 2 S, H 2, NH 4, CH 4), tanto como lo hacen hoy. Algunos respiraderos también son altos en metales como plomo, hierro, níquel, zinc cobre, etc. Cuando se combinan con sus sustratos arcillosos o corticales, estos minerales podrían haber proporcionado superficies catalíticas para mejorar la síntesis de moléculas orgánicas. ¿Estas condiciones localizadas podrían haber sido el foco de la experimentación química prebiótica que condujo a los orígenes de la vida? Veamos dos tipos de respiraderos hidrotermales de aguas profundas reconocidos hoy en día: volcánicos y alcalinos.
1. Orígenes en un respiradero hidrotermal de alto calor (ahumador negro)
La energía libre disponible de un respiradero hidrotermal volcánico provendría del alto calor (temperaturas que van a 350 o C) y los minerales y químicos expulsados del manto terrestre. A continuación se ilustra un respiradero hidrotermal volcánico.

Las condiciones asumidas para los respiraderos hidrotermales volcánicos prebióticos podrían haber soportado síntesis catalítica de moléculas orgánicas a partir de precursores inorgánicos (ver Volcanic Vents y formación de moléculas orgánicas). Los catalizadores habrían sido minerales metálicos (níquel, hierro, etc.). Las reacciones químicas probadas incluyen algunas que recuerdan a las reacciones bioquímicas en las células quimioautótrofas vivas hoy en día. Günter Wächtershäuser propuso la teoría mundial hierro-azufre de los orígenes de la vida en estos respiraderos, también llamados “fumadores negros”. Estos respiraderos ahora arrojan grandes cantidades de CH 4 y NH 4 y los experimentos favorecen la idea de que los agregados de hierro-azufre en y alrededor de los fumadores negros podrían proporcionar superficies catalíticas para la formación prebiótica de moléculas orgánicas como metanol y ácido fórmico a partir de CO disuelto 2 y los CH 4 y NH 4 procedentes de las rejillas de ventilación. Wächtershäuser también se le atribuye la idea de que la selección prebiótica actuó no tanto en reacciones químicas aisladas, sino en agregados de reacciones metabólicas.
Podríamos pensar en agregados metabólicos como vías bioquímicas o múltiples vías integradas. Wächtershäuser propuso la selección de reacciones químicas cíclicas que liberaban energía libre utilizable por otras reacciones. Esta evolución metabólica prebiótica de las químicas de reacción (más que una evolución química más simple) habría sido esencial para los orígenes de la vida. Una variedad de extremófilos (por ejemplo, arqueas termófilas) que ahora viven en y alrededor de los fumadores negros parece ser testimonio de los orígenes de la vida de los fumadores negros.
Si bien la idea de seleccionar vías metabólicas tiene gran mérito, existen problemas con un escenario de orígenes de vida en respiraderos hidrotermales volcánicos. Por un lado, sus altas temperaturas habrían destruido tantas moléculas orgánicas como se crearon. Además, las arqueas extremófilas que ahora se encuentran alrededor de estos respiraderos volcánicos no pueden ser descendientes directos de ninguna célula que pudiera haberse originado allí. La filogenia de Woese muestra claramente que las arqueas comparten un linaje con eucariotas (no eubacterias - ver arriba). Por lo tanto, la vida celular extremófila originada en los respiraderos debió haber dado lugar primero a un LUCA más moderado antes de luego morir de sí mismos, después de lo cual los extremófilos volverían a evolucionar independientemente para recolonizar los respiraderos! Esto mitiga frente a un escenario extremófilos- primeros orígenes. Ante estas preocupaciones, las propuestas recientes se centran en los orígenes de la vida en respiraderos hidrotermales alcalinos menos extremos.
2. Orígenes en una ventilación alcalina de aguas profundas (ahumador blanco)
De los diversos escenarios aquí discutidos, un origen de vida autotrófica en respiraderos alcalinos es una de las alternativas más satisfactorias a un origen soupy de células heterotróficas. Para empezar, a temperaturas cercanas a los 100 o C-150 o C, los respiraderos alcalinos (fumadores blancos) no son tan calientes como los fumadores negros. A continuación se muestra un respiradero alcalino.

Otras condiciones químicas y físicas de los respiraderos alcalinos también son consistentes con un escenario de origen de vida dependiente de la evolución metabólica. Por un lado, la interfaz de los respiraderos alcalinos con las aguas ácidas del océano tiene el potencial teórico de generar muchas moléculas orgánicas diferentes [Shock E, Canovas P. (2010) The potential for abiotic organic synthesis and biosynthesis at seas afloor hydrothermal systems. Geofluidos 10 (1-2) :161-92)]. En simulaciones de laboratorio de condiciones de ventilación alcalina, la presencia de CO 2 disuelto favorece la serpentinización, una reacción de agua y calor con serpentinita, mineral que contiene hierro que se encuentra en tierra y en la corteza oceánica. A continuación se muestra una muestra de serpentinita.

La serpentinización experimental produce hidrocarburos y una oxidación acuosa caliente del hierro produce H 2 que podría representar abundante H 2 en las emisiones actuales de los ahumadores blancos. También, durante la serpentinización, un mineral llamado olivino [(Mg +2, Fe +2) 2 SiO 4] reacciona con CO 2 disuelto para formar metano (CH 4). Entonces, la primera precondición de vida, la creación energéticamente favorable de moléculas orgánicas, es posible en respiraderos alcalinos.
Los defensores de los orígenes celulares en un océano no reductor del Hadeano tardío también se dieron cuenta de que las moléculas orgánicas formadas en un respiradero alcalino (o cualquier) se dispersarían y se neutralizarían rápidamente en las aguas ácidas de los océanos más amplios. De alguna manera, los orígenes en un planeta no reductor tuvieron que incluir alguna manera de contener moléculas orgánicas recién formadas desde el principio y potenciar una mayor evolución bioquímica. Entonces, ¿cuáles fueron las condiciones en un respiradero alcalino que podrían haber contenido moléculas orgánicas y conducir a la evolución metabólica y, en última instancia, a los orígenes de la vida? ¡Consideremos una propuesta intrigante que llegue a una respuesta!
La estructura de roca porosa de los respiraderos alcalinos actuales proporciona microespacios o microcompartimentos que podrían haber capturado líquidos alcalinos emitidos por los fumadores blancos. Resulta que las condiciones en los respiraderos alcalinos actuales también apoyan la formación de biopelículas de hidrocarburos. Los microcompartimentos revestidos con tales biopelículas podrían haber formado una membrana prebiótica primitiva contra una “pared celular” rocosa, dentro de la cual quedarían atrapadas las aguas alcalinas. El resultado sería un gradiente natural de protones entre las soluciones alcalinas de moléculas orgánicas atrapadas en los microcompartimentos y las aguas ácidas circundantes del océano. ¿Sucedió todo esto?
¡Quizás! Sin un ambiente rico en nutrientes, los heterótrofos primero no son una opción. Eso deja sólo la opción alternativa: un escenario de autótrofos primero para los orígenes de la vida. Nick Lane y sus compañeros de trabajo propusieron que los gradientes de protones eran la fuerza selectiva detrás de la evolución de las químicas metabólicas tempranas en el escenario de ventilación alcalina (Prebiotic Proton Gradient Energy Fuels Origins of Life). Organizadas alrededor de compartimentos de biopelículas, estructuras prebióticas y químicas habrían aprovechado la energía libre de los gradientes de protones naturales. En otras palabras, las primeras protocélulas, y luego las células, pueden haber sido quimioautótrofos.
Por último, pero no menos importante, ¿cómo podrían las químicas quimioautotróficas en un planeta no reductor haber apoyado la formación de polímeros, así como la replicación de polímeros? Hoy vemos el almacenamiento y replicación de información en ácidos nucleicos como separado de la catálisis enzimática de reacciones bioquímicas. Pero, ¿están todos tan separados? Si la replicación es la reproducción fiel de la información necesaria para una célula, entonces la catálisis enzimática asegura la producción redundante de todas las moléculas esenciales para hacer la célula! Dicho de otra manera, si la síntesis catalizada de polímeros es la replicación de las moléculas caballo de batalla que realizan tareas celulares, entonces lo que llamamos 'replicación' no es más que la replicación de la información de ácido nucleico necesaria para reproducir fielmente estas moléculas caballo de batalla. Entonces, ¿hubo una selección temprana, coordinada y concurrente de mecanismos para el metabolismo catalizado así como la síntesis y replicación catalizada de polímeros? Volveremos a estas preguntas en breve, cuando consideremos los orígenes de la vida en un mundo de ARN.
Los orígenes de la vida en una atmósfera no reductora (y libre de oxígeno) plantean preguntas adicionales. ¿Los gradientes de protones proporcionarían suficiente energía libre para alimentar y organizar los orígenes de la vida? Si es así, ¿cómo aprovecharon realmente las células derivadas del metabolismo quimiosmótico prebiótico la energía de un gradiente de protones? Antes de la vida, ¿las protocélulas ya eran capaces de transducir energía libre de gradiente en energía libre de químicos? ¿Y el ATP fue seleccionado para retener energía libre de químicos desde el principio? Alternativamente, ¿el alivio del gradiente se acopló al principio a la síntesis de otros compuestos intermedios de alta energía con, por ejemplo, enlaces tioéster? Más tarde, ¿cómo las células formadas en los respiraderos alcalinos escaparon de los respiraderos para colonizar el resto del planeta?
Independientemente de cómo se capturó inicialmente la energía libre de gradiente de protones, el LUCA quimioautotrófico ya debió haber estado usando bombas de protones unidas a membrana y una ATPasa para aprovechar la energía libre de gradiente para producir ATP, ya que todos sus descendientes lo hacen. Por último, ¿cuándo evolucionó la fotoautotrofia (específicamente fotoautotrofia oxigénica)? ¿Fue un evento evolutivo tardío? ¿Es posible que las células fotosintéticas evolucionaran bastante temprano entre algunos de los habitantes quimioautotróficos de los fumadores blancos, esperando su tiempo antes de explotar en escena para crear nuestro ambiente oxigénico?
3. Heterotrophs-First vs Autótrophs-First: Algunas Consideraciones Evolutivas
En el escenario de ventilación alcalina, el metabolismo quimiosmótico es anterior a la vida. Por lo tanto, las primeras células quimioautótrofas no necesitaron las reacciones fermentativas requeridas por las células en un escenario de origen heterótrofo primero. A pesar de que todas las células vivas hoy en día incorporan una forma de metabolismo glucolítico, la glucólisis puede no ser la vía bioquímica más antigua conocida, como hemos pensado durante tanto tiempo.
En apoyo de una evolución posterior de las enzimas glicolíticas, las de las arqueas muestran poco parecido estructural con las de las bacterias. Si la heterotrofia fermentativa fuera un desarrollo evolutivo tardío, LUCA y sus primeros descendientes carecerían de una vía glucolítica bien desarrollada. En cambio, el LUCA debe haber sido una de las muchas células autótrofas 'experimentales', muy probablemente un quimioautótrofo que deriva energía libre de químicos inorgánicos en el ambiente. Para dar cuenta de la heterotrofia en los tres dominios de la vida, debe haber evolucionado separadamente en las dos ramas antecedentes que descienden del último ancestro común universal de organismos bacterianos, arqueales y eucariotas. La filogenia que se muestra a continuación ilustra el escenario de autótrofos primero.
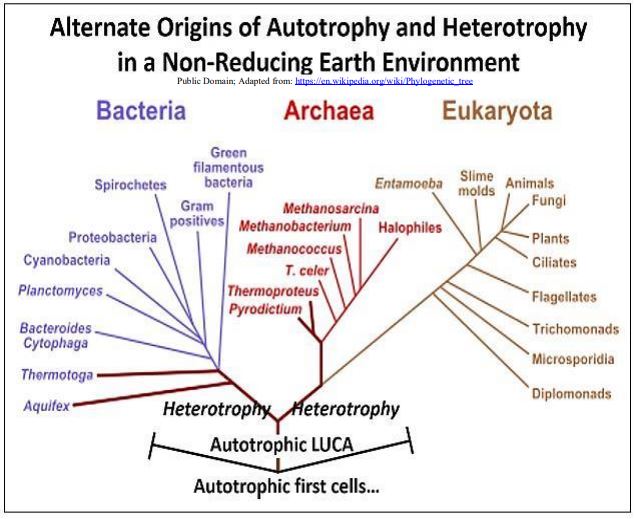
4. Resumiendo
La especulación sobre los orígenes de la vida comienza tratando de identificar una fuente de energía libre con la que hacer moléculas orgánicas. Las primeras células podrían haber sido heterótrofos formados en un ambiente terrestre reductor, a partir del cual los autótrofos evolucionaron posteriormente. Por otro lado, las primeras células pueden haber sido autótrofas formadas en condiciones no reductoras en ausencia de una sopa primordial. Entonces, solo después de que estos autótrofos hubieran producido suficiente energía libre de nutrientes para sostenerlos emergieron tardíamente heterótrofos. Descubrimientos que sugieren que la atmósfera terrestre era una no reductora más que hace 4 mil millones de años (poco después de la formación del planeta), y que había vida en la tierra hace 3.95 mil millones de años favorecen los orígenes metabólicos de la vida autótrofica en un respiradero térmico, probablemente un respiradero alcalino. Sin embargo, quedan dudas sobre los orígenes de la vida en condiciones no reductoras. Incluso la composición de la atmósfera prebiótica todavía está en disputa (ver Tierra no reductora- ¡No tan rápido! ).
Por ahora, dejemos de lado estas preocupaciones por un momento y volvamos a los acontecimientos que nos llevan desde el LUCA y sus primeros descendientes a las elaboradas químicas comunes a todas las células hoy en día. Las descripciones que siguen son conjeturas educadas sobre los caminos tomados desde el principio hacia la celularidad familiar ahora en la tierra. Abordan principalmente la selección de mecanismos catalíticos, el metabolismo replicativo, la red de vías bioquímicas que se cruzan y la comunicación química aún más intrincada que organizó la función celular y la complejidad.
352 Soporte filogenético para los autótrofos: primeros orígenes de la vida