
10.7: Traducción eucariota
- Page ID
- 53375
La traducción eucariota, al igual que con la transcripción, es satisfactamente similar (desde el punto de vista de un estudiante que estudia, o desde uno de conservación evolutiva) al caso procariótico. El proceso de iniciación es un poco más complicado, pero los procesos de elongación y terminación son los mismos, pero con homólogos eucariotas de los factores de elongación y liberación apropiados.
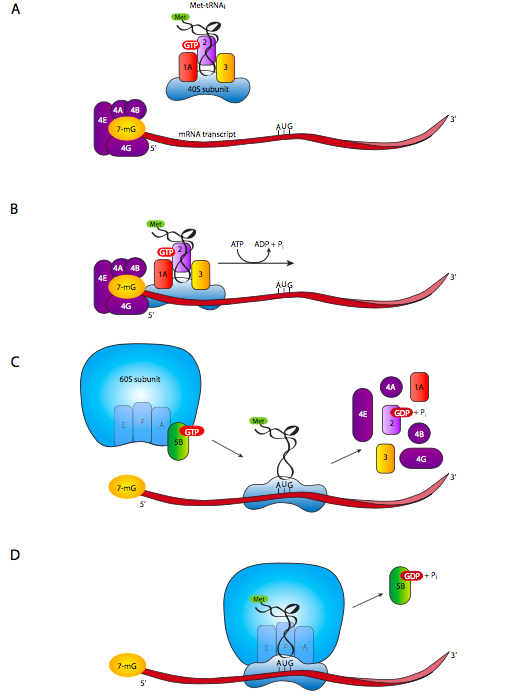
Para los eucariotas, cada ARNm codifica uno y solo un gen (a diferencia de los transcritos multigénicos como los operones), por lo que no hay mucha duda de qué AUG es un codón de inicio, y cuáles son solo metioninas regulares. Por lo tanto, no se requiere una secuencia Shine-Dalgarno en eucariotas. La subunidad ribosómica pequeña, acompañada de los factores de iniciación eucariotas eIF-3, eIF-2 y Met-ARNt i, juntos conocidos como el complejo terneario, se une a eIF-1a. Mientras tanto, eIF-4A, -4B, -4E y -4G se unen a la tapa 5' (7-metiguanosina) del ARNm (g. 7A). El complejo de subunidad pequeña y el complejo eIF4/ ARNm de unión a la tapa interactúan para formar el complejo 43S, que luego comienza a escanear el ARNm de 5' a 3' buscando el primer AUG.
Por lo general, pero no siempre, el primer AUG es el codón de inicio para los genes eucariotas. Sin embargo, el contexto del AUG importa, y es un codón de inicio mucho más fuerte (es decir, más frecuentemente reconocido y usado) si hay un residuo de purina (A o G) en -3 y un G en +4. Ver Kozak, M., Biochimie 76: 815-821, 1994.
Una vez que el complejo de escaneo 43S ha encontrado el codón de inicio, los factores de iniciación caen y llega la subunidad ribosómica grande. La subunidad ribosómica grande se ha unido a eIF-6, lo que impide que se reasocie con subunidades pequeñas, y su eliminación se requiere primero. Otro factor, eIF-5 entra en escena durante el proceso de acoplamiento entre las subunidades ribosómicas grandes y pequeñas, y se requiere la hidrólisis de un GTP unido a eIF-5 para completar el acoplamiento de las subunidades y la formación de un ribosoma funcional completo en el ARNm.
El alargamiento es funcionalmente el mismo que en los procariotas excepto que las funciones de EF-Tu son atendidas por EF-1a, también con hidrólisis de GTP. EF-2 es el análogo eucariota de EF-G, y utiliza la hidrólisis de GTP para la translocación del ribosoma. La terminación usa homólogos eucariotas de los factores de liberación, aunque ERF-1 toma el lugar tanto de RF-1 como de RF-2 procariotas.
Aunque se pueden formar poliribosomas (también conocidos como polisomas) tanto en ARNm procariotas como eucariotas, los polisomas eucariotas tienen una torsión adicional. Técnicamente, un polisoma es simplemente un ARNm con múltiples ribosomas que lo traducen simultáneamente, pero en eucariotas, el polisoma también tiene una morfología única porque utiliza PABPI, o proteína de unión a poli-A. Esta proteína no solo se une a la cola 3' poli-A de un ARNm, sino que también interactúa con los factores de iniciación de eIF-4, que de esta manera enrolla el ARNm en una forma circular. De esa manera, una vez que el ribosoma llega al final del gen y se libera del ARNm, se encuentra físicamente cerca del comienzo del ARNm para comenzar a traducir de nuevo.