
14.1: Introducción a la Transducción de Señales
- Page ID
- 53420

Existen tres modos primarios de comunicación intercelular. Estos son
- contacto directo entre moléculas de señalización unidas a las membranas de dos células adyacentes,
- señales solubles de corto alcance que se difunden a distancias cortas, y
- señales solubles de largo alcance que se secretan en la circulación para ser transportadas a cualquier parte del cuerpo.
Un ejemplo de señalización yuxtacrina se ejemplifica por la actividad de algunas proteínas de adhesión celular o ECM, como la laminina, que no solo permiten que una célula se mueva sobre ellas, sino que actúan como señales para promover un aumento de la motilidad. Esto probablemente sucede por la activación de los receptores de integrina en la célula en movimiento, que luego inician y coordinan cambios a través del resto de la célula para lograr el cambio en la actividad. Otro ejemplo es la vía Delta-Notch utilizada en patrones embrionarios.
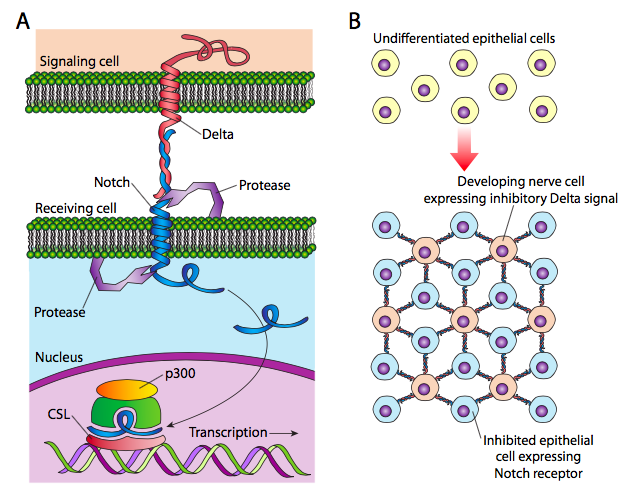
Delta, una proteína transmembrana en la célula de señalización, se une a Notch, un receptor en la célula receptora. Notch altera su conformación, permitiendo que su dominio citoplásmico sea cortado por la g-secretasa. El dominio citoplásmico luego se transloca al núcleo, donde actúa como factor de transcripción activante al unirse con CSL. En el ejemplo esbozado en la Figura\(\PageIndex{1}\) B, la regulación positiva estocástica de delta en una célula activa la muesca en las células circundantes, que luego activa una ruta de diferenciación específica para ellas. Así, la célula central puede ser una neurona sensorial, como una célula pilosa, mientras que las que la rodean inmediatamente son células de soporte como la glía. Este tipo de señalización impone un patrón de espaciamiento en la expresión de neuronas (u otra célula).
La vía Delta-Notch está bien caracterizada y algo más complicada que la retratada en el párrafo anterior. La escisión de Notch involucra dos proteasas y dos sitios. Una vez que el dominio citoplásmico Notch se une a CSL, desplaza una serie de co-represores unidos al CSL, y también recluta MAM (Mastermind-1) como coactivador. MAM recluta histona acetilasas para permitir aumentar aún más la transcripción de genes dirigidos, pero también recluta quinasas que inician el proceso de apuntar al dominio citoplásmico de Notch para la destrucción mediada por ubiquitina. La expresión de genes controlados por Notch es así autorregulada y se apaga poco después de que Delta ya no esté disponible. Opinión escrita en R.A. Kovall, Curr. Opin. Struct. Biol. 17: 117-27, 2007.
Las señales de difusión limitada de vecinos cercanos se llama señalización paracrina, y algunas veces las señales pueden actuar sobre los receptores justo en la célula que secretó la señal, que sería señalización autocrina. Las señales paracrinas solo están activas si pueden unirse a una célula por encima de una concentración crítica para activar una vía de señalización. Por lo tanto, a medida que las señales se difunden lejos de la fuente, hay un corte, más allá del cual la concentración de señal es insuficiente para activar una célula receptora. Los factores de crecimiento suelen ser señales paracrinas. Aunque a menudo fomentan el crecimiento, también suelen ser factores de supervivencia. En ese contexto, el Factor de Crecimiento Nervioso (NGF) es secretado por las células diana que luego recompensan a las neuronas que hacen las conexiones correctas al proporcionar NGF para su supervivencia. Aquellas neuronas que se dirigen en la dirección equivocada, son incapaces de obtener NGF, y no sobreviven, promoviendo la eficiencia y una mejor relación señal:ruido dentro del sistema nervioso.
La señalización endocrina es esencialmente señalización de todo el cuerpo. Una señal producida por una glándula productora de hormonas se secreta en el torrente sanguíneo, donde se vuelve accesible a casi cualquier célula del cuerpo. Por supuesto, no todas las células responderán a la hormona: como cualquier otro caso de señalización intercelular, la respuesta depende totalmente de los receptores, por lo que solo reaccionarán las células que tengan receptores para reconocer la señal. Por ejemplo, el estrógeno se libera en la circulación, pero en las hembras, solo algunos órganos muestran un impacto significativo cuando los niveles de estrógeno se alteran significativamente. La mayoría de los tejidos no se ven afectados. Las señales endocrinas pueden circular en otros fluidos extracelulares como la linfa.