
14.5: Señalización de iones de calcio
- Page ID
- 53410

La señalización mediante el aumento del calcio citosólico es un mecanismo de coordinación intracelular importante y ubicuo. Ya vimos que se requiere la liberación de Ca 2+ en las células musculares para permitir la contracción de cada sarcómero, y el posicionamiento del retículo sarcoplásmico hace posible cambios rápidos en la concentración casi simultáneos en toda la célula. Otro mecanismo fisiológico extremadamente importante que se basa en el calcio es la fertilización. Inmediatamente después de la penetración del óvulo por el esperma, una ola de aumento intracelular de Ca 2+ se extiende a través del óvulo a partir del punto de fertilización. Esto activa CaMKII (una quinasa) y calcineurina (una fosfatasa). Ambos son necesarios para superar la detención meiótica y también pueden ser necesarios en el desarrollo embrionario inicial mediante el control de la descondensación de la cromatina, la formación de la envoltura nuclear y el movimiento y fusión de los dos núcleos.
El CaMKII mencionado anteriormente es la quinasa II dependiente de Ca 2+ /calmodulina, e ilustra un tema bastante común, que es el uso de proteínas de unión a Ca 2+ como activadores de detección intermedios de Ca 2+. La calmodulina es una proteína ubicua de unión al calcio en eucariotas y su importancia se destaca por la extraordinariamente alta homología entre especies. En los niveles citosólicos normales de Ca 2+, la afinidad relativamente baja de los 4 sitios de unión de Ca 2+ en la calmodulina están sin rellenar. Pero cuando las concentraciones de Ca 2+ aumentan, ocupan los sitios, provocando un cambio conformacional en la calmodulina y permitiendo que interactúe con otras proteínas. Además de la calmodulina, la troponina-C y la PKC, algunas otras proteínas importantes sensibles al calcio son calsequestrina, una proteína tampón Ca 2+, gelsolina, la enzima seccionadora de f-actina, la proteasa calpaína y calretinina, un activador de la guanilciclasa (que hace el segundo mensajero cGMP).
La guanilato (guanilil) ciclasa también es un actor importante en la transducción de señales por óxido nítrico (NO). El óxido nítrico es un gas producido por la acción de la óxido nítrico sintasa (NOS) sobre el sustrato aminoácido arginina. Se utiliza como una señal súper soluble que pasa a través de las células fácilmente. Sin embargo, requiere concentraciones relativamente altas para el efecto fisiológico, por lo que es estrictamente un factor paracrino que trabaja en vecinos cercanos. Quizás el ejemplo mejor estudiado de la señalización de NO es la vasodilatación, en la que las células endoteliales que expresan NOS de un vaso sanguíneo liberan NO a las células del músculo liso que las rodean. El NO se une y estimula la guanilato ciclasa. El aumento resultante en la concentración de GMPc conduce a la relajación a través de múltiples dianas de proteína quinasa G.
Sildena l (Viagra) y sus hermanos químicos aprovechan esta vía inhibiendo fosfodiesterasas específicas de GMPc (PDE5) que normalmente descomponen las GMPc para limitar la respuesta al NO. Sin embargo, cabe señalar que aunque la expresión de PDE5 es limitada, se expresa no solo en los genitales sino también en la retina.
Finalmente, ninguna discusión sobre la transducción de señales estaría completa sin al menos una mención fugaz de la extraordinaria diafonía (Figura\(\PageIndex{12}\)) que puede ocurrir entre las diferentes vías mencionadas.
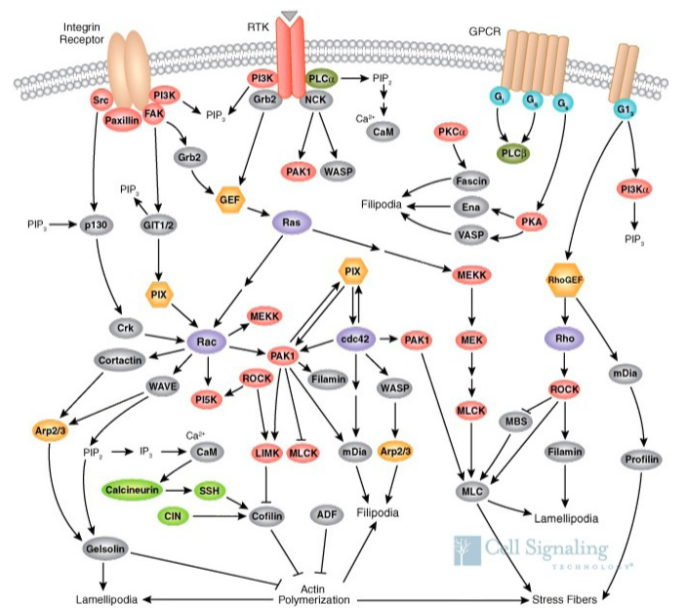
La Figura representa solo una pequeña parte de la señalización que ocurre dentro de una celda en movimiento. No solo algunas partes de la célula forman filopodios para ayudar a determinar a dónde ir, otras partes o albordear la lamelipodia, y otras, inducen a las proteínas motoras a reorganizar el citoesqueleto de la manera adecuada para facilitar el transporte a granel internamente incluso cuando el borde de ataque de la célula está impulsando reenviar para hacer contactos externamente. Todo esto debe coordinarse por diafonía entre sistemas de señalización como se representa, sin mencionar la señalización relacionada con el metabolismo, o la expresión génica, o incluso el ciclo celular, todos los cuales están sucediendo simultáneamente dentro de la célula.