4.6: Firmas génicas de microARN (miARN)
- Page ID
- 54771

Un ejemplo de regiones genómicas funcionales sujetas a altos niveles de conservación son las secuencias que codifican microARN (miARN). Los miARN son moléculas de ARN que se unen a secuencias complementarias en la región 3' no traducida de moléculas de ARNm dirigidas, causando silenciamiento génico. ¿Cómo encontramos las firmas evolutivas para los genes de miARN y sus dianas, y podemos utilizarlas para obtener nuevos conocimientos sobre sus funciones biológicas? Veremos que esta es una tarea desafiante, ya que los miARN dejan una señal evolutiva altamente conservada pero muy sutil.
Desafío Computacional
Predecir la ubicación de los genes de miARN y sus objetivos es un problema computacionalmente desafiante. Podemos buscar regiones “horquilla”, donde encontramos secuencias de nucleótidos que son complementarias entre sí y predicen una estructura de horquilla. Pero de 760,355 horquillas similares a miARN que se encuentran en la célula, solo 60-100 eran verdaderos miARN. Entonces, para hacer cualquier prueba que nos dé regiones estadísticamente probables de ser miARN, necesitamos una prueba con 99.99% de especificidad.
La Figura 4.25 es un ejemplo del patrón de conservación para genes de miARN. Se pueden observar las dos estructuras en horquilla conservadas en las regiones roja y azul, con una región de baja conservación en el medio. Este patrón es característico de los miARN.

Al analizar las características evolutivas y estructurales específicas del miARN, podemos usar combinaciones de estas características para seleccionar regiones de miARN con un enriquecimiento >4.500 veces en comparación con horquillas aleatorias. Los siguientes son ejemplos de características que ayudan a seleccionar los miARN:
- Los miARN se unen a motivos diana altamente conservados en la UTR 3'
- Los miARN se pueden encontrar en intrones de genes conocidos
Figura 4.26: Nuevo miARN en intron© fuente desconocida. Todos los derechos reservados. Este contenido está excluido de nuestra licencia Creative Commons. Para obtener más información, consulte http://ocw.mit.edu/help/faq-fair-use/.
- Los miARN tienen preferencia por la cadena positiva del ADN y por los factores de transcripción
- Los miARN normalmente no se encuentran en elementos exónicos y repetitivos del genoma (contraejemplo en la Figura 4.29).
- Los nuevos miARN pueden agruparse con miARN conocidos, especialmente si están en la misma familia o tienen un origen común

Estas características de las regiones codificadoras de miARN se pueden agrupar en familias estructurales, lo que permite construir clasificadores basados en ARN conocidos en cada familia. Las consideraciones energéticas para la estructura del ARN pueden ser utilizadas para apoyar esta clasificación en familias. Dentro de cada familia, se produce la conservación ortóloga (genes en diferentes especies para una misma función con el gen ancestral común) y la conservación paráloga (genes duplicados dentro de una misma especie que evolucionaron para servir a diferentes funciones).
Evolutiva
- Correlación con el perfil de conservación
- MFE del pliegue de consenso
- Índice de conservación de la estructura
Estructural
- Estabilidad de horquilla (puntuación z MFE)
- Número de bucles asimétricos
- Número de bucles simétricos
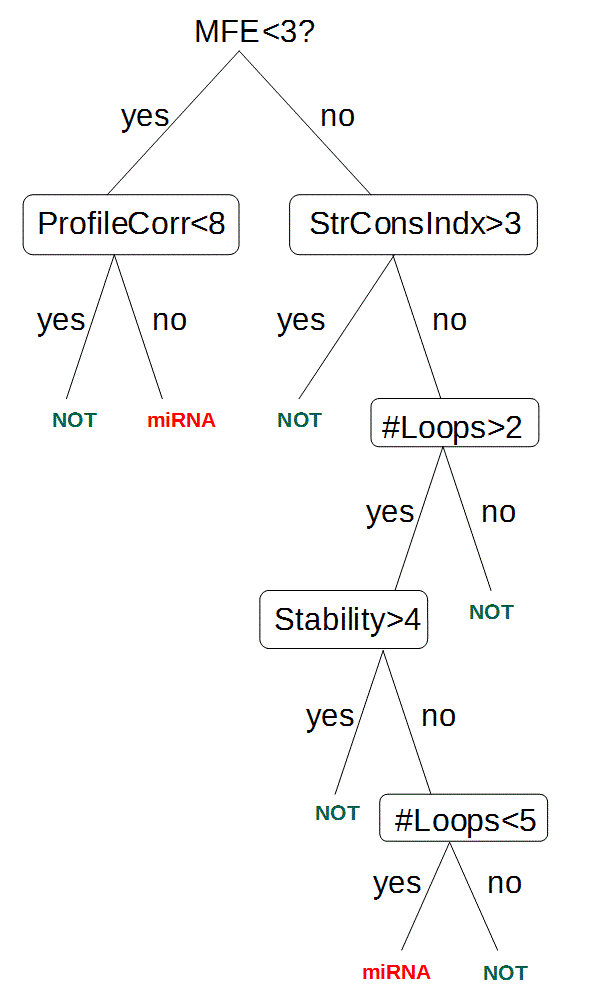
Podemos combinar varias características en una prueba usando un árbol de decisión, como se ilustra en la Figura 4.28. En cada nodo del árbol, se aplica una prueba que determina qué rama se seguirá a continuación. El árbol se recorre comenzando desde la raíz hasta que se alcanza un nodo terminal, momento en el que el árbol emitirá una clasificación. Un árbol de decisión puede ser entrenado usando un cuerpo de subsecuencias genómicas clasificadas, después de lo cual se puede usar para predecir si las nuevas subsecuencias son miARN o no. Además, muchos árboles de decisión se pueden combinar en un “bosque aleatorio”, donde se entrenan varios árboles de decisión. Cuando se necesita clasificar una nueva secuencia de nucleótidos, cada árbol vota sobre si es o no un miARN, y luego se agregan los votos para determinar la clasificación final.
La aplicación de esta técnica al genoma de la mosca mostró 101 horquillas por encima del punto de corte de 0.95, redescubriendo 60 de 74 miARN conocidos, prediciendo 24 nuevos miARN que fueron validados experimentalmente, y encontrando 17 candidatos adicionales que mostraron evidencia de diversa función.
Genes de miARN inusuales
Se encontraron las siguientes cuatro “sorpresas” al observar genes específicos de miARN:
Sorpresa 1 Ambas hebras podrían ser expresadas y funcionales. Por ejemplo, en el gen miR-Iab-4, la expresión de las cadenas sentido y antisentido se ve en distintos dominios embrionarios. Ambas hebras puntúan > 0.95 para la predicción de miARN.
Sorpresa 2 Algunos miARN pueden tener múltiples extremos 5' para un solo brazo de miARN, lo que da evidencia de un sitio de inicio impreciso. Esto podría dar lugar a múltiples productos maduros, cada uno potencialmente con sus propios objetivos funcionales.
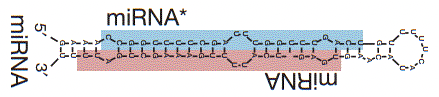
Sorpresa 3 Las regiones de miRNA* de alta puntuación (el brazo estelar es complementario a la secuencia real de miARN) están muy altamente expresadas, dando lugar a regiones del genoma que están altamente expresadas y contienen elementos funcionales.
Sorpresa 4 Se ha demostrado que tanto miR—10 como miR-10* son reguladores Hox muy importantes, lo que lleva a la predicción de que los miRNAs podrían ser “reguladores maestros de Hox”. Las páginas 10 y 11 del primer conjunto de conferencias 5 diapositivas muestran la importancia de los miARN que forman una red de regulación para diferentes genes Hox.
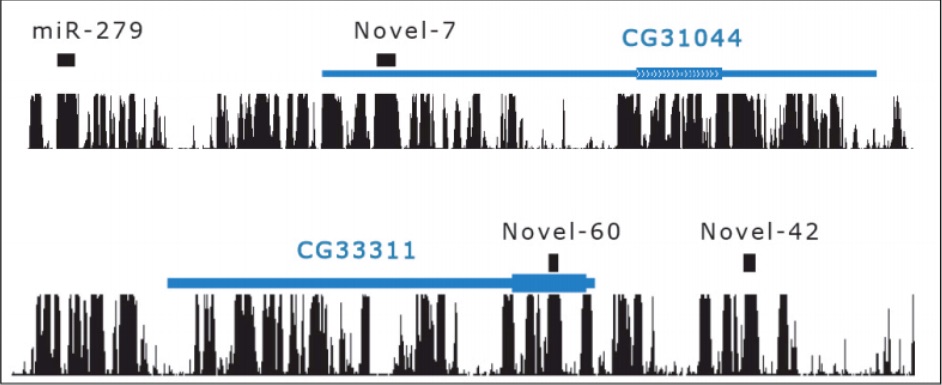
Ejemplo: Reexamen de genes codificadores de proteínas 'dudosos'
Dos genes, CG31044 y CG33311 fueron rechazados independientemente porque sus patrones de conservación no coincidieron con los característicos de las firmas evolutivas de una proteína (ver Sección 4.5). Se identificaron como miARN precursores con base en propiedades genómicas y altos niveles de expresión (Lin et al.). Este es un raro ejemplo de miARN que se encuentra en secuencias previamente exónicas e ilustra el desafío de identificar firmas evolutivas de miARN.