
3.2: División y Movimiento Celular: Escote - Gastrulación de Rana y Labio Dorsal del Blastopore
- Page ID
- 53093

Escote
Las etapas de escisión de la embriogénesis suelen ser muy similares entre los animales. Durante la escisión, un embrión de una sola célula se divide rápidamente para formar una bola de células, llamada blástula. Esta bola puede ser hueca por dentro, tener una región hueca o ser bastante sólida. Las etapas tempranas de escisión son diferentes de las divisiones celulares posteriores en que producen rápidamente muchas células de aspecto muy similar. Si los morfógenos se distribuyen asimétricamente en el huevo, entonces estas células pueden diferenciarse mediante especificación autónoma incluso en etapas tempranas. La especificación condicional generalmente comienza a ocurrir hacia el final de las etapas de escisión cuando la división celular se ralentiza y las células comienzan a comunicarse más extensamente entre sí. A veces esto se llama la “transición de blástula media” o MBT. El MBT demarca la transición entre el uso principalmente de células maternas y determinantes del destino (ARN y proteína) al embrión produciendo sus propios ARN y proteínas. Un experimento interesante que muestra el alcance de esta transición es bloquear la transcripción usando los fármacos alfa-amanitina o actinomicina D, dos inhibidores de la acción de la ARN polimerasa. Si estos fármacos se aplican a una etapa temprana de escisión, las escisiones embrionarias proceden normalmente, lo que sugiere que los procesos de desarrollo y la división celular en esta etapa temprana están utilizando factores derivados de la madre ya presentes en el embrión. Si los fármacos se aplican a embriones en etapas posteriores, el desarrollo se detiene, lo que sugiere que en este punto los transcritos derivados cigóticos son necesarios para el desarrollo y la división celular. El tiempo de la MBT y la clasificación de proteínas y ARN maternos en diferentes células en etapa de escisión varía ampliamente entre diferentes animales.
Lee más detalles sobre Escote aquí, enfócate en cualquier sección que te parezca interesante y sobre todo figura 8.5. www.ncbi.nlm.nih.gov/libros/nbk9992/ #_A1678_
Gastrulación
La gastrulación se refiere a un tiempo de gran flujo en embriones animales. Durante la gastrulación las células producidas durante la escisión comienzan a moverse entre sí. A medida que se mueven, cambian su entorno local y entran en contacto con nuevas celdas. Este cambio en el entorno local significa que las células pueden a) estar expuestas a diferentes morfógenos extracelulares y b) hacer contacto con nuevos compañeros de señalización célula-célula. El movimiento de las células (Figura 3) sobre (epibolía), debajo (involución), entre (intercalación), internamente (invaginación) o alejándose de una lámina epitelial (ingreso y delaminación) crea capas de tejido. A medida que las células que comprenden estas capas de tejido se encuentran con nuevos morfógenos y nuevos compañeros de señalización, se especifican como capas germinales.
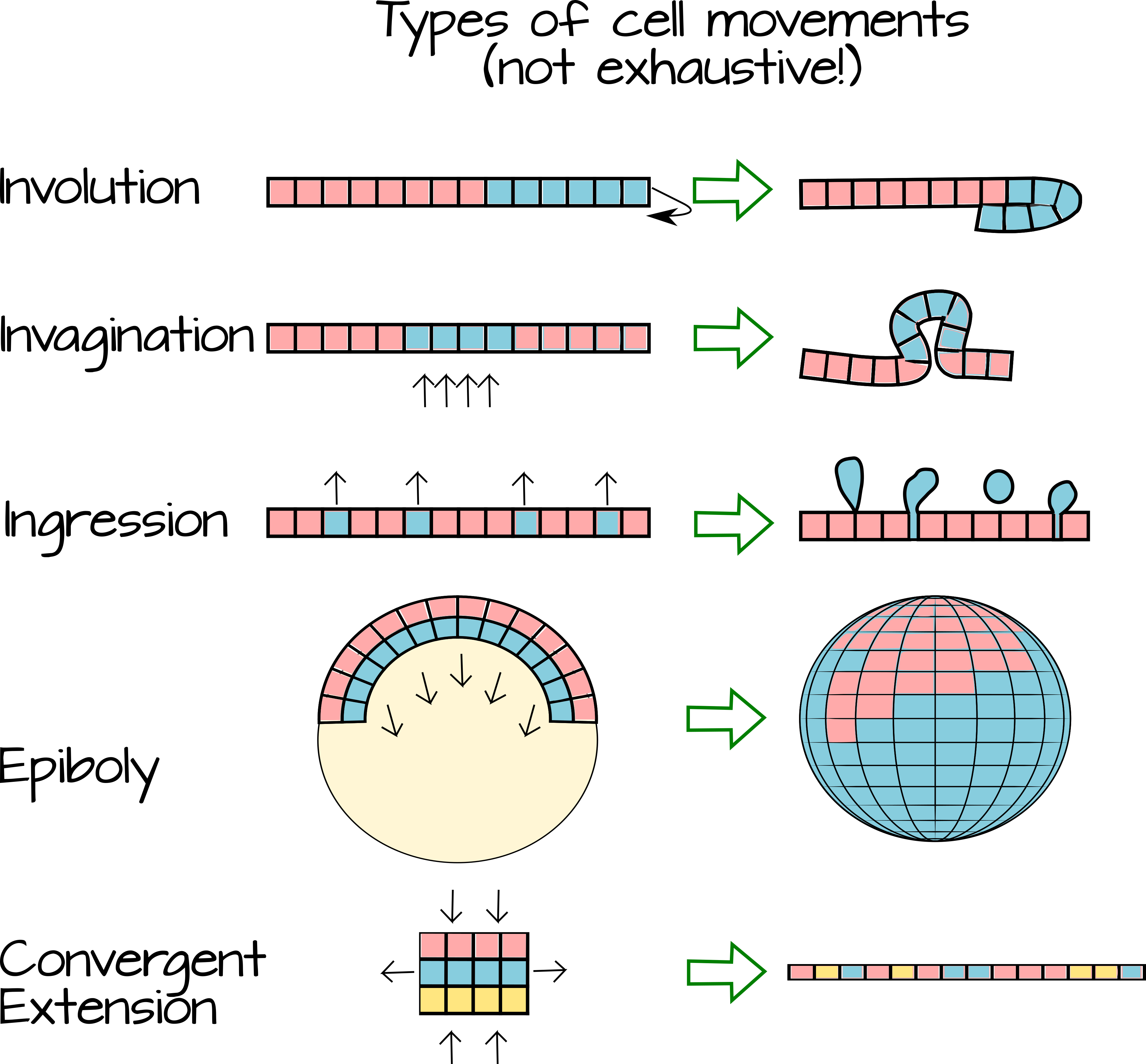
El siguiente video muestra un embrión de rana viva sometido a escisión y gastrulación y también muestra una visión interna animada de cómo las células se mueven dentro del embrión. Tenga en cuenta los diferentes tipos de movimientos. https://www.youtube.com/watch?v=riSA1mo86Kg
Cambio de Socios
Como se mencionó anteriormente, la gastrulación realiza dos funciones clave en un embrión animal en desarrollo. Primero, crea capas de tejido (capas germinales) a partir de un conjunto de células bastante homogéneas. En segundo lugar, permite nuevas interacciones entre las células y coloca a las células en nuevas posiciones morfogénicas. A medida que las células cambian a sus parejas con el tiempo, se vuelven destinadas a un camino específico. Por ejemplo, compare una imagen de un mapa de destino de una rana en etapa de blastula con el mapa de destino de una rana gastrulada (Figura 4).
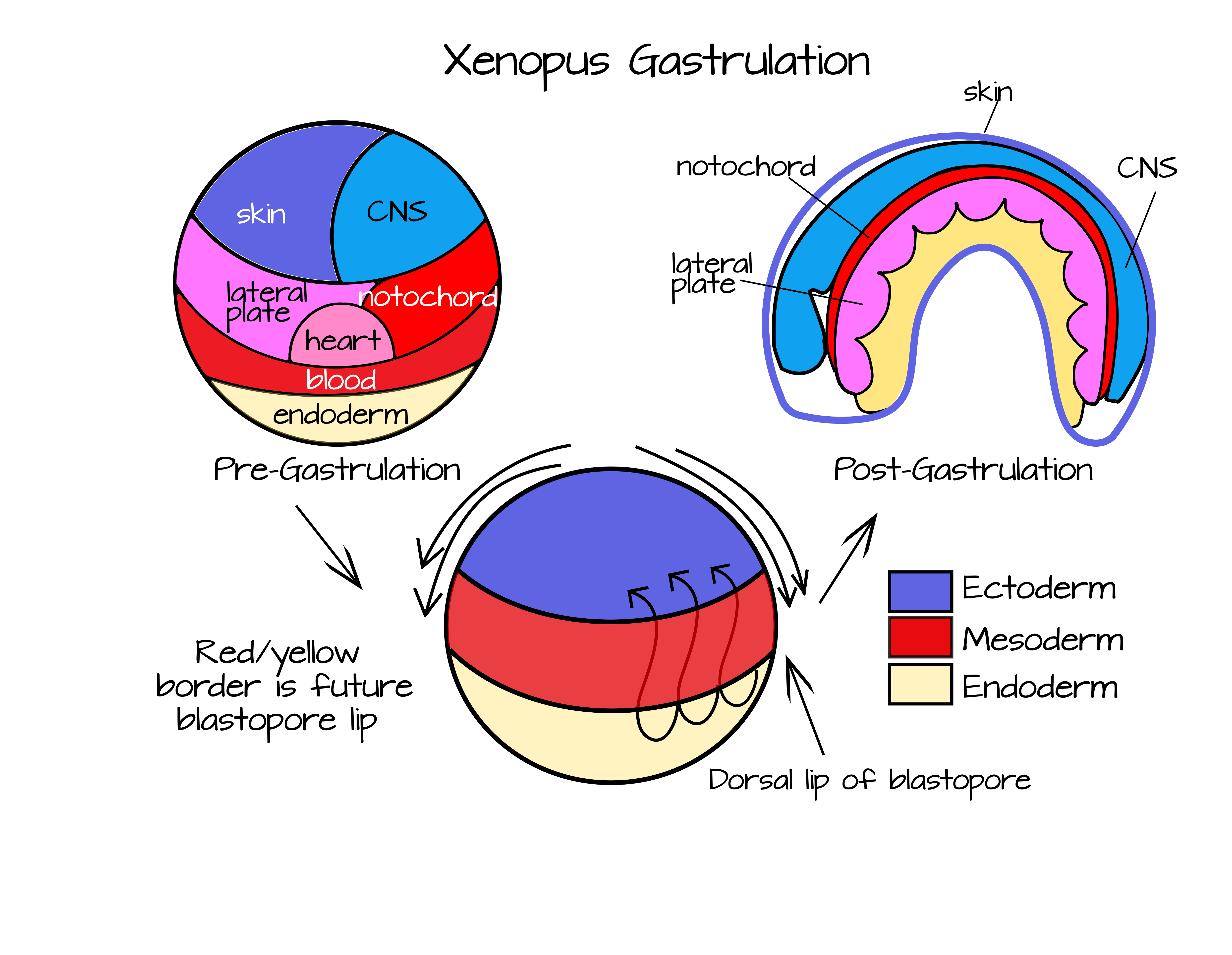
Mientras que la rana en estadio blástula parece casi aleatoria en su mapeo, la rana gastrulada tiene todos sus tejidos/órganos en posiciones más o menos adultas. La etapa de la blástula aún no ha experimentado movimientos celulares extensos y las células están en gran parte sin especificar. Si las células se eliminan durante la etapa de blástula, otras células a menudo podrán asumir el destino de las células faltantes. Por ejemplo, los gemelos idénticos se desarrollan a partir de un solo embrión que se ha “dividido” en dos después de múltiples divisiones celulares. En lugar de que cada mitad del embrión haga solo la mitad de un animal cuando se rompe el contacto entre las dos masas celulares, cada una de ellas se convierte en un animal completo. Uno de mis conjuntos favoritos de experimentos de todos los tiempos, los experimentos del organizador Spemann-Mangold, muestra hasta qué punto la comunicación célula-célula puede gobernar el desarrollo. Antes de discutir eso me gustaría resumir brevemente la gastrulación de ranas (sin ofender a otros animales, pero son mis gastruladores favoritos). Una mirada más profunda a la gastrulación de la rana se puede encontrar aquí: www.ncbi.nlm.nih.gov/books/nbk10113/ y un mapa de destino interactivo para Xenopus está disponible en https://www.xenbase.org/anatomy/static/xenbasefate.jsp
Gastrulación de rana y labio dorsal del Blastopore
Después de la fertilización y rotación cortical, el embrión de rana se divide aproximadamente en cuatro cuadrantes. Estos están delimitados por los dos ejes principales A/V (animal-vegetal) y D/V (dorsoventral). Todavía no existe un eje anteroposterior, este eje se generará durante la gastrulación. https://www.youtube.com/watch?v=EPMgHMnwW28
En el transcurso de la gastrulación, el área roja (mesodermo futuro) en la Figura 5 se involucionará bajo el área azul (ectodermo futuro) y se extenderá para formar una capa en el techo del blastocoel (Figuras 3 y 4). Esta involución ocurre más extensamente en el lado dorsal del embrión (el lado opuesto a la entrada de espermatozoides), pero también ocurre a lo largo del resto del “labio blastoporo”. Para visualizar el labio del blastóporo, abra bien la boca. Tus labios representan mesodermo y el resto de tu rostro representa ectodermo (¡perdón no endodermo en este!). Usa tus labios para cubrirte los dientes. Esto es físicamente similar a la involución. Ahora imagina que tus labios podrían adelgazarse y alargarse y cubrir el interior de tu boca. Es así como las células mesodérmicas terminan como una segunda capa bajo el ectodermo. Por último, empieza a cerrar la boca. Esto es similar a la epibolía, cuando las células ectodérmicas se mueven sobre la superficie del embrión para cubrir el mesodermo y el endodermo.
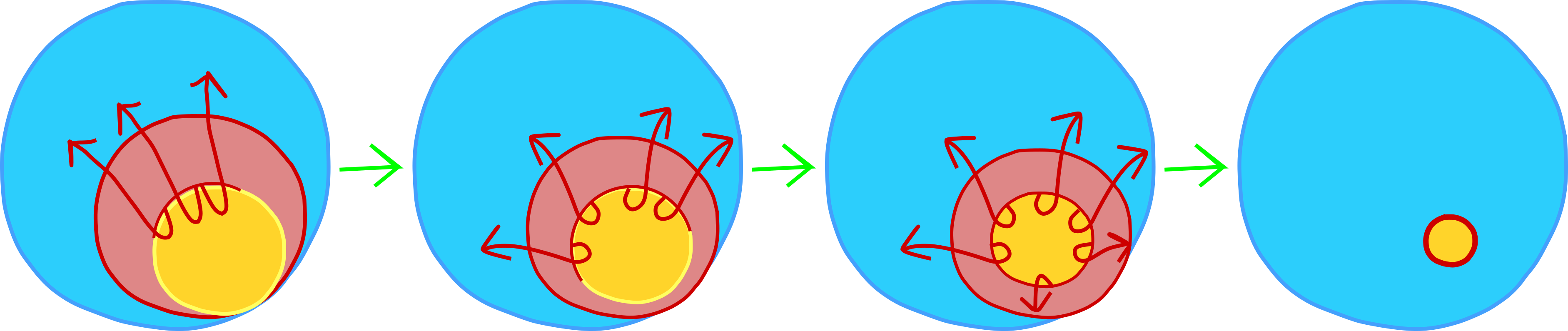
Para entender el papel del organizador de Spenmann-Mangold, es necesario tener en cuenta estos movimientos. En particular, hay que pensar en el labio dorsal del blastoporo, el lugar donde primero comienza la involución y ocurre más extensamente. Si miras el mapa del destino de la Figura 4, notarás que el mesodermo dorsal (el mesodermo en el labio dorsal del blastoporo) se convertirá en la notocorda. Estas células son las primeras mesodérmicas en internalizarse durante la epibolía. Este bloque de células se alargará y adelgazará para convertirse en una varilla de tejido mesodérmico que abarca la longitud del embrión. Esto correrá a lo largo del eje anteroposterior, al igual que tu columna vertebral corre a lo largo de tu eje anterioposterior. Adicionalmente, las señales proteicas de la notocorda ayudarán a modelar el resto del embrión, como descubrieron Hans Spemann e Hilde Mangold.