
15.11.4: Potenciación a Largo Plazo (LTP)
- Page ID
- 56926

Todo aprendizaje depende de la memoria. La formación de memorias parece ocurrir en dos fases separadas, la primera memoria a corto plazo. (e.g., Los seres humanos sometidos a un tratamiento de electroshock (para aliviar su depresión) no pueden recordar eventos que ocurrieron justo antes del tratamiento, pero su memoria de eventos anteriores no se ve afectada) que siguieron por la formación de memoria a largo plazo. El daño a los lóbulos temporales del cerebro puede resultar en la pérdida de la capacidad de recordar nuevos aprendizajes durante más de aproximadamente una hora. Dos sistemas que han sido particularmente útiles para elaborar las bases celulares y moleculares para la formación de la memoria son la sensibilización en la babosa marina Aplysia y el estudio de la potenciación a largo plazo (LTP).
Potenciación a largo plazo (LTP)
Las ratas y los ratones pueden ser entrenados para resolver tareas simples. Por ejemplo, si un ratón se coloca en un charco de agua turbia, nadará alrededor hasta que encuentre una plataforma oculta para subir. Con la repetición, el ratón pronto aprende a ubicar la plataforma más rápidamente. Presumiblemente lo hace con la ayuda de señales visuales colocadas alrededor del perímetro de la alberca porque no puede ver ni oler la plataforma misma.
Las ratas o ratones que han tenido una parte de su cerebro llamada hipocampo dañada, no pueden aprender esta tarea, aunque continúan resolviéndola rápidamente si fueron entrenados antes de su daño cerebral. Esto sugiere que las neuronas en el hipocampo son necesarias para este tipo de aprendizaje. A diferencia del resto del cerebro, se producen nuevas neuronas en el hipocampo a lo largo de la vida. Surgen de un charco de células madre en el cerebro. La integración de neuronas recién formadas en los circuitos existentes del hipocampo facilita el aprendizaje de nuevos recuerdos (así como el olvido de los antiguos).
Demostrar potenciación a largo
El comportamiento de ciertas sinapsis en la región “CA1" del hipocampo de la rata (o ratón) es consistente con que son esenciales para esta forma de memoria a largo plazo. Rebanadas del hipocampo pueden ser removidas y sus neuronas CA1 estudiadas in vitro con electrodos de registro. La estimulación rápida e intensa de las neuronas presinápticas evoca potenciales de acción en la neurona postsináptica. Esto es justo lo que esperaríamos de las propiedades de las sinapsis.
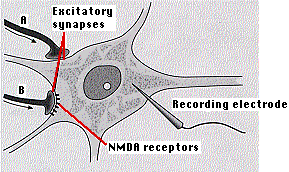
Lo que es notable de este sistema es que con el tiempo estas sinapsis se vuelven cada vez más sensibles de manera que un nivel constante de estimulación presináptica se convierte en una mayor salida postsináptica (gráfica a la derecha). Esta potenciación a largo plazo puede durar semanas. El tratamiento de una porción de hipocampo con un medicamento llamado ácido aminofosfonovalérico (“APV”) bloquea la formación de LTP. Esto se debe a que el APV bloquea la acción de los receptores NMDA, un subconjunto de receptores postsinápticos que normalmente responden al neurotransmisor excitador glutamato (Glu). Los receptores NMDA (sinapsis B anterior) se distinguen de otros receptores activados por Glu-por ser estimulados por el fármaco, N-metil-D-aspartato (NMDA).
Los receptores NMDA contienen un canal transmembrana que permite la difusión facilitada de iones de calcio (Ca 2+) a través de la membrana plasmática de la sinapsis. La unión de Glu (o NMDA) y D-serina liberada de un astrocito cercano a estos receptores abre el canal permitiendo que el Ca 2+ fluya en si, y solo si, la misma célula postsináptica ha sido despolarizada simultáneamente por otras sinapsis en ella (sinapsis A anterior). (El dibujo está enormemente simplificado: cada neurona CA1 tiene decenas de miles de sinapsis en ella).
La afluencia de Ca 2+ a la neurona activa una enzima llamada quinasa II dependiente de calcio-calmodulina (CaMKII). Las quinasas unen grupos fosfato a las proteínas y, al hacerlo, alteran su funcionamiento. En este caso, CaMKII fosforila un segundo tipo de receptor Glu llamado receptores AMPA, lo que los hace más permeables a los iones de sodio (Na +) disminuyendo así el potencial de reposo de la célula y haciéndola más sensible a los impulsos entrantes. Además, hay evidencia de que la actividad de CaMKII incrementa el número de receptores AMPA en la sinapsis.
La capacidad de hacer ratones transgénicos ha proporcionado herramientas para probar este modelo de LTP.
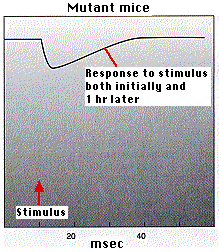
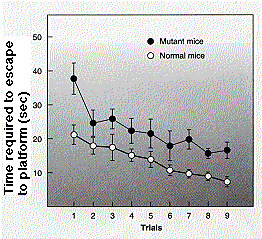
Los ratones que son homocigotos para un transgén CaMKII mutante no logran desarrollar LTP. Esto se demostró (por A. J. Silver, et. al., en Science 257: 206, 1992) de dos maneras:
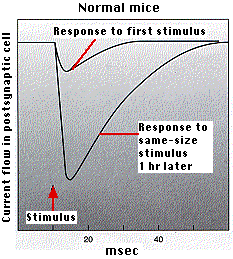
- midiendo la corriente en la célula postsináptica de ratones normales y ratones mutantes. La gráfica anterior (izquierda) muestra que los ratones mutantes no desarrollan el aumento en el flujo de corriente que hacen los ratones normales (gráfico anterior).
- El mismo fallo de LTP ocurre cuando los ratones se prueban en la plataforma oculta (gráfico a la derecha).
Los ratones transgénicos que producen cantidades extra grandes de receptores de NMDA muestran LTP potenciada como se muestra por
- mayores corrientes postsinápticas en su hipocampo
- su rendimiento mejorado en la prueba de plataforma oculta
- y rendimiento mejorado en otras pruebas de aprendizaje y memoria
Estos hallazgos fueron reportados en la edición del 2 de septiembre de 1999 de Nature por Tang, Y-P, et al. Los experimentos descritos anteriormente muestran que las manipulaciones que afectan la respuesta eléctrica postsináptica (EPSP) de las neuronas estimuladas eléctricamente también afectan el comportamiento aprendido. No muestran que el aprendizaje induce aumento de EPSP en neuronas postsinápticas del hipocampo. Ahora, los investigadores del MIT han hecho precisamente eso.
Utilizaron ratas en las que implantaron una matriz de electrodos de grabación muy espaciados en la región CA1 del hipocampo. Estas ratas fueron luego colocadas en un aparato de entrenamiento donde aprendieron en un solo ensayo que pasar de una cámara iluminada a otra oscura les daría un shock.
En solo 30 minutos algunos, pero nunca todos, de los electrodos de registro captaron EPSP incrementaron en las neuronas CA1 y el número de receptores AMPA aumentó en las células CA1. Entonces, el aprendizaje de esta respuesta condicionada produjo los cambios eléctricos y sinápticos de LTP pero sólo en ciertas regiones del hipocampo. Presumiblemente otros tipos de aprendizaje producirían LTP en otras partes de la región CA1. (Se puede leer el reporte de su trabajo en Whitlock, J. R., et al. , Ciencia, 25 de agosto de 2006.)
LTP Temprano vs LTP Tardío
LTP ocurre en dos fases:
- uno temprano (en la primera hora más o menos) que implica una mayor sensibilidad de la sinapsis sin que se produzca ninguna transcripción de genes nuevos o traducción de ARNm
- una tardía que requiere transcripción de genes nuevos y traducción de ARNm y da como resultado un aumento en el número de receptores AMPA acompañado de un aumento en el tamaño de la conexión sináptica. Estos cambios persisten por muchas horas e incluso muchos días. Sin embargo, el aumento de la formación del receptor AMPA parece requerir estimulación continua porque (en ratas, al menos) interferir con el proceso borra la LTP tardía (y la memoria) incluso un mes después.
Esto es aún más evidencia de que los recuerdos se adquieren en dos fases; temprana y tardía. La LTP tardía puede implicar no solo la adición de receptores AMPA a las sinapsis existentes, sino la formación de sinapsis completamente nuevas. Investigaciones en Ginebra, Suiza han demostrado que la formación de LTP en cerebros de ratas coincide con la formación de sinapsis adicionales (al menos una más) entre el axón presináptico terminal y la dendrita con la que sinapsis. (Informe de Toni, N., et al, Nature, 25 Nov 99). Presumiblemente esto, también, aumenta la eficiencia de la transmisión sináptica.
Resumen
- La estimulación rápida e intensa de neuronas CA1 en el hipocampo las despolariza.
- La unión de Glu y D-serina a sus receptores NMDA los abre.
- Los iones Ca 2+ fluyen hacia la célula a través de los receptores NMDA y se unen a la calmodulina.
- Esto activa la quinasa II dependiente de calcio-calmodulina (CaMKII).
- CaMKII fosforila los receptores AMPA haciéndolos más permeables a la entrada de iones Na + y aumentando así la sensibilidad de la célula a la despolarización.
- Con el tiempo CaMKII también aumenta el número de receptores AMPA en la sinapsis.
- El aumento de la expresión génica (es decir, síntesis de proteínas, quizás de los receptores AMPA) también ocurre durante el desarrollo de LTP.
- El agrandamiento de las conexiones sinápticas y tal vez la formación de sinapsis adicionales ocurren durante la formación de LTP.
También se ha demostrado LTP en neuronas del cerebelo.
Depresión a Largo Plazo (LTD)
La estimulación eléctrica lenta y débil de las neuronas CA1 también provoca cambios a largo plazo en las sinapsis, en este caso, una reducción en su sensibilidad. Esto se llama depresión a largo plazo o LTD. Reduce el número de receptores AMPA en la sinapsis. La depresión a largo plazo también ocurre en preparaciones aisladas de neuronas de la babosa marina, Aplysia y el cerebelo de ratones durante el desarrollo de una respuesta condicionada (CR)