
4.5: Conexiones a otras vías metabólicas
- Page ID
- 54344

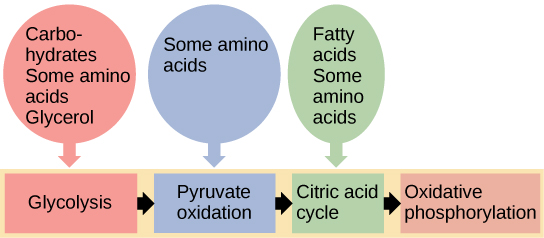
Has aprendido sobre el catabolismo de la glucosa, que aporta energía a las células vivas. Pero los seres vivos consumen más que solo glucosa como alimento. ¿Cómo un sándwich de pavo, que contiene proteínas, proporciona energía a tus células? Esto sucede porque todas las vías catabólicas para carbohidratos, proteínas y lípidos eventualmente se conectan con la glucólisis y las vías del ciclo del ácido cítrico (Figura\(\PageIndex{1}\)). Las vías metabólicas deben considerarse porosas, es decir, las sustancias ingresan por otras vías y otras sustancias salen por otras vías. Estas vías no son sistemas cerrados. Muchos de los productos en una ruta particular son reactivos en otras vías.
Conexiones de otros azúcares al metabolismo de la glucosa
El glucógeno, un polímero de glucosa, es una molécula de almacenamiento de energía a corto plazo en animales. Cuando hay ATP adecuado presente, el exceso de glucosa se convierte en glucógeno para su almacenamiento. El glucógeno se elabora y se almacena en el hígado y el músculo. El glucógeno se extraerá del almacenamiento si bajan los niveles de azúcar en la sangre. La presencia de glucógeno en las células musculares como fuente de glucosa permite que el ATP se produzca por más tiempo durante el ejercicio.
La sacarosa es un disacárido hecho de glucosa y fructosa unidos entre sí. La sacarosa se descompone en el intestino delgado, y la glucosa y la fructosa se absorben por separado. La fructosa es uno de los tres monosacáridos de la dieta, junto con la glucosa y la galactosa (que forma parte del azúcar de la leche, el disacárido lactosa), que se absorben directamente en el torrente sanguíneo durante la digestión. El catabolismo tanto de fructosa como de galactosa produce el mismo número de moléculas de ATP que la glucosa.
Conexiones de las proteínas al metabolismo de la glucosa
Las proteínas son descompuestas por una variedad de enzimas en las células. La mayoría de las veces, los aminoácidos se reciclan en nuevas proteínas. Si hay exceso de aminoácidos, sin embargo, o si el cuerpo está en estado de hambruna, algunos aminoácidos se derivarán hacia vías de catabolismo de glucosa. Cada aminoácido debe tener su grupo amino eliminado antes de entrar en estas vías. El grupo amino se convierte en amoníaco. En mamíferos, el hígado sintetiza urea a partir de dos moléculas de amoníaco y una molécula de dióxido de carbono. Por lo tanto, la urea es el principal producto de desecho en los mamíferos a partir del nitrógeno que se origina en los aminoácidos, y deja el cuerpo en la orina.
Conexiones de los lípidos al metabolismo de la glucosa
Los lípidos que están conectados a las vías de la glucosa son el colesterol y los triglicéridos. El colesterol es un lípido que contribuye a la flexibilidad de la membrana celular y es un precursor de las hormonas esteroides. La síntesis de colesterol comienza con acetil CoA y procede en una sola dirección. El proceso no se puede revertir, y no se produce ATP.
Los triglicéridos son una forma de almacenamiento de energía a largo plazo en animales. Los triglicéridos almacenan aproximadamente el doble de energía que los carbohidratos. Los triglicéridos están hechos de glicerol y tres ácidos grasos. Los animales pueden hacer la mayoría de los ácidos grasos que necesitan. Los triglicéridos pueden elaborarse y descomponerse a través de partes de las vías del catabolismo de la glucosa. El glicerol se puede fosforilar y procede a través de la glucólisis. Los ácidos grasos se rompen en unidades de dos carbonos que entran en el ciclo del ácido cítrico.
EVOLUCIÓN EN ACCIÓN: Vías de Fotosíntesis y Metabolismo
La fotosíntesis y el metabolismo celular constan de varias vías muy complejas. Generalmente se piensa que las primeras células surgieron en un ambiente acuoso, una “sopa” de nutrientes. Si estas células se reproducían con éxito y sus números subían de manera constante, se deduce que las células comenzarían a agotar los nutrientes del medio en el que vivían, ya que desplazaban los nutrientes hacia sus propias células. Esta situación hipotética habría resultado en la selección natural favoreciendo a aquellos organismos que podrían existir mediante el uso de los nutrientes que permanecieron en su entorno y al manipular estos nutrientes en materiales que podrían utilizar para sobrevivir. Adicionalmente, la selección favorecería a aquellos organismos que pudieran extraer el valor máximo de los nutrientes disponibles.
Se desarrolló una forma temprana de fotosíntesis que aprovechó la energía del sol utilizando compuestos distintos al agua como fuente de átomos de hidrógeno, pero esta vía no produjo oxígeno libre. Se piensa que la glucólisis se desarrolló antes de este tiempo y podría aprovechar la producción de azúcares simples, pero estas reacciones no fueron capaces de extraer completamente la energía almacenada en los carbohidratos. Una forma posterior de fotosíntesis utilizó el agua como fuente de iones de hidrógeno y generó oxígeno libre. Con el tiempo, la atmósfera se oxigenó. Los seres vivos se adaptaron para explotar esta nueva atmósfera y permitieron que la respiración tal y como la conocemos evolucionara. Cuando se desarrolló el proceso completo de fotosíntesis tal como la conocemos y la atmósfera se oxigenó, las células finalmente pudieron utilizar el oxígeno expulsado por la fotosíntesis para extraer más energía de las moléculas de azúcar utilizando el ciclo del ácido cítrico.
Resumen
La descomposición y síntesis de carbohidratos, proteínas y lípidos se conectan con las vías del catabolismo de la glucosa. Los carbohidratos que también pueden alimentar el catabolismo de la glucosa incluyen galactosa, fructosa y glucógeno. Estos se conectan con la glucólisis. Los aminoácidos de las proteínas se conectan con el catabolismo de la glucosa a través de piruvato, acetil CoA y componentes del ciclo del ácido cítrico. La síntesis de colesterol comienza con acetil CoA, y los componentes de los triglicéridos son recogidos por acetil CoA y entran en el ciclo del ácido cítrico.