
6.4: División celular procariota
- Page ID
- 53918

Procariotas como las bacterias se propagan por fisión binaria. Para los organismos unicelulares, la división celular es el único método para producir nuevos individuos. Tanto en células procariotas como eucariotas, el resultado de la reproducción celular es un par de células hijas que son genéticamente idénticas a la célula parental. En los organismos unicelulares, las células hijas son individuos.
Para lograr el resultado de células hijas idénticas, algunos pasos son esenciales. El ADN genómico debe replicarse y luego asignarse a las células hijas; los contenidos citoplásmicos también deben dividirse para dar a ambas nuevas células la maquinaria para sostener la vida. En las células bacterianas, el genoma consiste en un único cromosoma de ADN circular; por lo tanto, se simplifica el proceso de división celular. La mitosis es innecesaria porque no hay núcleo ni múltiples cromosomas. Este tipo de división celular se llama fisión binaria.
Fsión binaria
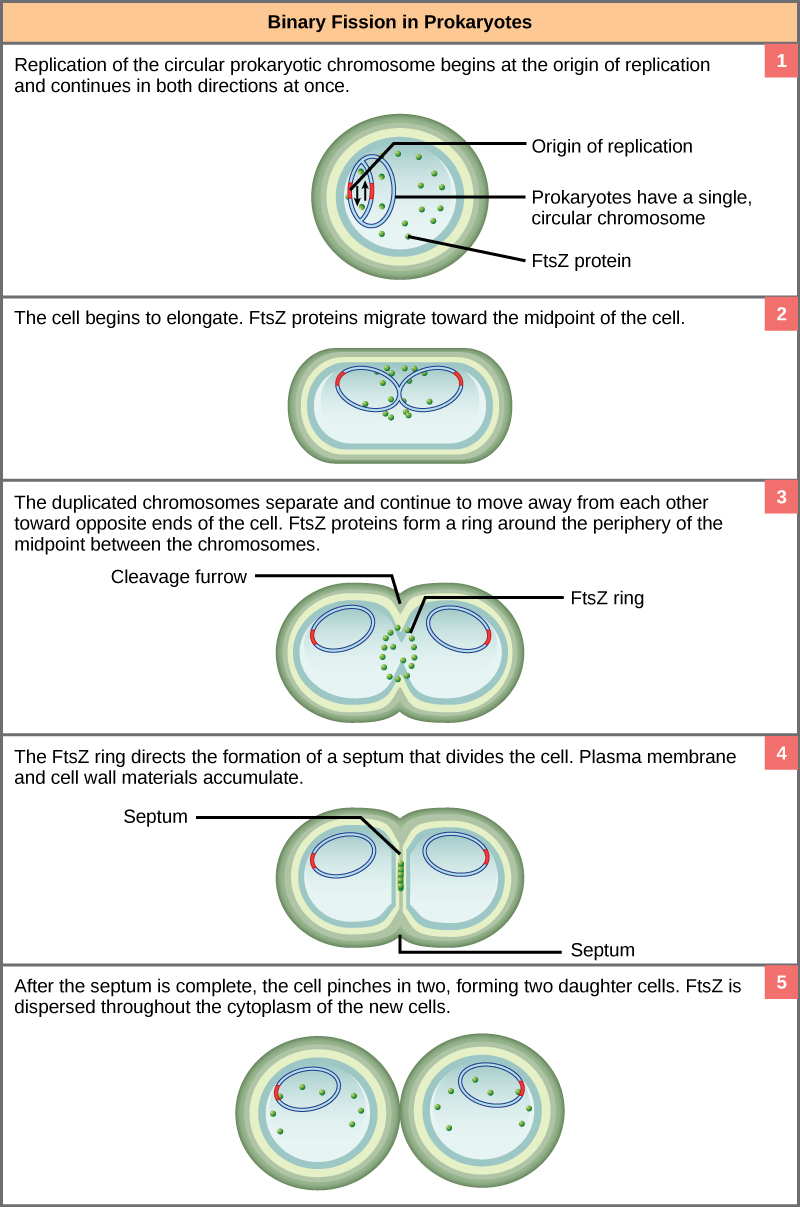
El proceso de división celular de los procariotas, llamado fisión binaria, es un proceso menos complicado y mucho más rápido que la división celular en eucariotas. Debido a la velocidad de la división celular bacteriana, las poblaciones de bacterias pueden crecer muy rápidamente. El único cromosoma circular de ADN de las bacterias no está encerrado en un núcleo, sino que ocupa una ubicación específica, el nucleoide, dentro de la célula. Al igual que en los eucariotas, el ADN del nucleoide está asociado con proteínas que ayudan a empaquetar la molécula en un tamaño compacto. Las proteínas de empaquetamiento de bacterias están, sin embargo, relacionadas con algunas de las proteínas involucradas en la compactación cromosómica de eucariotas.
El punto de partida de replicación, el origen, está cerca del sitio de unión del cromosoma a la membrana plasmática (Figura\(\PageIndex{1}\)). La replicación del ADN es bidireccional, alejándose del origen en ambas cadenas del bucle de ADN simultáneamente. A medida que se forman las nuevas hebras dobles, cada punto de origen se aleja de la unión de la pared celular hacia los extremos opuestos de la celda. A medida que la célula se alarga, la membrana creciente ayuda en el transporte de los cromosomas. Después de que los cromosomas hayan despejado el punto medio de la célula alargada, comienza la separación citoplasmática. Se forma un tabique entre los nucleoides desde la periferia hacia el centro de la célula. Cuando las nuevas paredes celulares están en su lugar, las celdas hijas se separan.
EVOLUCIÓN EN ACCIÓN: Aparato Huso Mitótico
El momento preciso y la formación del huso mitótico es fundamental para el éxito de la división celular eucariota. Las células procariotas, por otro lado, no sufren mitosis y por lo tanto no tienen necesidad de un huso mitótico. Sin embargo, la proteína ftsZ que juega un papel tan vital en la citocinesis procariota es estructural y funcionalmente muy similar a la tubulina, el bloque de construcción de los microtúbulos que componen las fibras del huso mitótico que son necesarias para los eucariotas. La formación de un anillo compuesto por unidades repetitivas de una proteína llamada FtsZ dirige la partición entre los nucleoides en procariotas. La formación del anillo ftsZ desencadena la acumulación de otras proteínas que trabajan juntas para reclutar nuevos materiales de membrana y pared celular en el sitio. Las proteínas FtsZ pueden formar filamentos, anillos y otras estructuras tridimensionales que se asemejan a la forma en que la tubulina forma microtúbulos, centriolos y diversos componentes del citoesqueleto. Además, tanto FtsZ como tubulina emplean la misma fuente de energía, GTP (guanosina trifosfato), para ensamblar y desmontar rápidamente estructuras complejas.
FtsZ y tubulina son un ejemplo de homología, estructuras derivadas de los mismos orígenes evolutivos. En este ejemplo, se presume que FtsZ es similar a la proteína ancestro tanto a la ftsZ moderna como a la tubulina. Si bien ambas proteínas se encuentran en organismos existentes, la función de la tubulina ha evolucionado y diversificado enormemente desde la evolución de su origen procariótico similar al FTSZ. Un estudio de la maquinaria de división celular en eucariotas unicelulares actuales revela pasos intermedios cruciales para la compleja maquinaria mitótica de eucariotas multicelulares (Tabla\(\PageIndex{1}\)).
| Estructura del material genético | División de material nuclear | Separación de células hijas | |
|---|---|---|---|
| Procariotas | No hay núcleo. El único cromosoma circular existe en una región del citoplasma llamada nucleoide. | Se produce a través de fisión binaria. A medida que se replica el cromosoma, las dos copias se mueven a extremos opuestos de la célula por un mecanismo desconocido. | Las proteínas FtsZ se ensamblan en un anillo que pellizca la célula en dos. |
| Algunos protistas | Los cromosomas lineales existen en el núcleo. | Los cromosomas se adhieren a la envoltura nuclear, la cual permanece intacta. El huso mitótico pasa a través de la envoltura y alarga la célula. No existen centriolos. | Los microfilamentos forman un surco de escisión que pellizca la célula en dos. |
| Otros protistas | Los cromosomas lineales existen en el núcleo. | Un huso mitótico se forma a partir de los centriolos y pasa a través de la membrana nuclear, que permanece intacta. Los cromosomas se adhieren al huso mitótico. El huso mitótico separa los cromosomas y alarga la célula. | Los microfilamentos forman un surco de escisión que pellizca la célula en dos. |
| Células animales | Los cromosomas lineales existen en el núcleo. | Se forma un huso mitótico a partir de los centriolos. La envoltura nuclear se disuelve. Los cromosomas se adhieren al huso mitótico, lo que los separa y alarga la célula. | Los microfilamentos forman un surco de escisión que pellizca la célula en dos. |
Resumen
Tanto en la división celular procariota como en la eucariota, el ADN genómico se replica y cada copia se asigna a una célula hija. Los contenidos citoplásmicos también se dividen de manera uniforme a las nuevas células. Sin embargo, existen muchas diferencias entre la división celular procariota y eucariota. Las bacterias tienen un solo cromosoma de ADN circular y no tienen núcleo. Por lo tanto, la mitosis no es necesaria en la división celular bacteriana. La citocinesis bacteriana es dirigida por un anillo compuesto por una proteína llamada FtsZ. El crecimiento interno de la membrana y el material de la pared celular desde la periferia de las células resulta en un tabique que finalmente forma las paredes celulares separadas de las células hijas.
Glosario
- fisión binaria
- el proceso de división celular procariota
- FtsZ
- un componente proteico similar a tubulina del citoesqueleto procariota que es importante en la citocinesis procariota (nombre origen: F ilamenting t emperature- s ensitive mutant Z)
- origen
- la región del cromosoma procariota en la que comienza la replicación
- tabique
- una pared formada entre células hijas bacterianas como precursor de la separación celular