
7.1: Reproducción Sexual
- Page ID
- 54115

La reproducción sexual fue una innovación evolutiva temprana después de la aparición de células eucariotas. El hecho de que la mayoría de los eucariotas se reproducen sexualmente es evidencia de su éxito evolutivo. En muchos animales, es el único modo de reproducción. Y sin embargo, los científicos reconocen algunas desventajas reales para la reproducción sexual. En la superficie, las crías que son genéticamente idénticas al progenitor pueden parecer más ventajosas. Si el organismo progenitor está ocupando exitosamente un hábitat, la descendencia con los mismos rasgos tendría un éxito similar. También existe el beneficio obvio para un organismo que puede producir descendencia por gemación asexual, fragmentación o huevos asexuales. Estos métodos de reproducción no requieren de otro organismo del sexo opuesto. No hay necesidad de gastar energía encontrando o atrayendo a una pareja. Esa energía se puede gastar en producir más descendencia. En efecto, algunos organismos que llevan un estilo de vida solitario han conservado la capacidad de reproducirse asexualmente. Además, las poblaciones asexuales sólo tienen individuos femeninos, por lo que cada individuo es capaz de reproducirse. En contraste, los machos en poblaciones sexuales (la mitad de la población) no están produciendo descendencia ellos mismos. Debido a esto, una población asexual puede crecer dos veces más rápido que una población sexual en teoría. Esto quiere decir que en competencia, la población asexual tendría la ventaja. Todas estas ventajas para la reproducción asexual, que también son desventajas para la reproducción sexual, deberían significar que el número de especies con reproducción asexual debería ser más común.
Sin embargo, los organismos multicelulares que dependen exclusivamente de la reproducción asexual son extremadamente raros. ¿Por qué es tan común la reproducción sexual? Esta es una de las preguntas importantes en biología y ha sido el foco de mucha investigación desde la segunda mitad del siglo XX hasta la actualidad. Una explicación probable es que la variación que crea la reproducción sexual entre las crías es muy importante para la supervivencia y reproducción de esas crías. La única fuente de variación en los organismos asexuales es la mutación. Esta es la última fuente de variación en los organismos sexuales. Además, esas diferentes mutaciones se reorganizan continuamente de una generación a otra cuando diferentes padres combinan sus genomas únicos, y los genes se mezclan en diferentes combinaciones por el proceso de meiosis. La meiosis es la división de los contenidos del núcleo que divide los cromosomas entre los gametos. La variación se introduce durante la meiosis, así como cuando los gametos se combinan en la fertilización.
EVOLUCIÓN EN ACCIÓN: La hipótesis de la reina
No cabe duda de que la reproducción sexual proporciona ventajas evolutivas a los organismos que emplean este mecanismo para producir descendencia. La pregunta problemática es ¿por qué, incluso ante condiciones bastante estables, la reproducción sexual persiste cuando es más difícil y produce menos descendencia para organismos individuales? La variación es el resultado de la reproducción sexual, pero ¿por qué son necesarias las variaciones continuas? Entra en la hipótesis de la Reina Roja, propuesta por primera vez por Leigh Van Valen en 1973. 1 El concepto fue nombrado en referencia a la carrera de la Reina Roja en el libro de Lewis Carroll, Through the Looking-Glass, en el que la Reina Roja dice que uno debe correr a toda velocidad solo para quedarse donde está uno.
Todas las especies coevolucionan con otros organismos. Por ejemplo, los depredadores coevolucionan con sus presas, y los parásitos coevolucionan con sus huéspedes. Un ejemplo notable de coevolución entre los depredadores y sus presas es la coadaptación única de los murciélagos voladores nocturnos y su presa de polilla. Los murciélagos encuentran a su presa emitiendo clics de tono alto, pero las polillas han evolucionado oídos simples para escuchar estos clics para que puedan evitar a los murciélagos. Las polillas también tienen comportamientos adaptados, como volar lejos del murciélago cuando lo escuchan por primera vez, o caer repentinamente al suelo cuando el murciélago está sobre ellas. Los murciélagos han evolucionado clics “silenciosos” en un intento de evadir la audición de la polilla. Algunas polillas han evolucionado la capacidad de responder a los clics de los murciélagos con sus propios clics como una estrategia para confundir las habilidades de ecolocación de los murciélagos.
Cada pequeña ventaja obtenida por la variación favorable le da a una especie una ventaja sobre competidores cercanos, depredadores, parásitos o incluso presas. El único método que permitirá que una especie en coevolución mantenga su propia parte de los recursos es también mejorar continuamente su capacidad de sobrevivir y producir descendencia. A medida que una especie gana ventaja, otras especies también deben desarrollar una ventaja o serán superadas. Ninguna especie avanza demasiado por delante porque la variación genética entre la progenie de la reproducción sexual proporciona a todas las especies un mecanismo para producir individuos adaptados. Las especies cuyos individuos no pueden mantenerse al día se extinguen. El frase de la Reina Roja era: “Se necesita toda la carrera que puedas hacer para quedarte en el mismo lugar”. Esta es una descripción acertada de la coevolución entre especies competidoras.
Ciclos de Vida de Organismos de Reproducción Sexual
La fertilización y la meiosis se alternan en los ciclos de vida sexual. Lo que ocurra entre estos dos eventos depende del organismo. El proceso de meiosis reduce a la mitad el número de cromosomas del gameto resultante. La fertilización, la unión de dos gametos haploides, restaura la condición diploide. Existen tres categorías principales de ciclos de vida en los organismos multicelulares: diploide-dominante, en la que la etapa diploide multicelular es la etapa de vida más obvia (y no hay etapa haploide multicelular), como ocurre con la mayoría de los animales incluyendo los humanos; haploide-dominante, en la que la etapa haploide multicelular se encuentra la etapa de vida más obvia (y no hay etapa diploide multicelular), como ocurre con todos los hongos y algunas algas; y alternancia de generaciones, en las que las dos etapas, haploide y diploide, son evidentes en un grado u otro dependiendo del grupo, como ocurre con las plantas y algunas algas.
Casi todos los animales emplean una estrategia de ciclo de vida diploide dominante en la que las únicas células haploides producidas por el organismo son los gametos. Los gametos se producen a partir de células germinales diploides, una línea celular especial que solo produce gametos. Una vez que se forman los gametos haploides, pierden la capacidad de volver a dividirse. No hay etapa de vida haploide multicelular. La fertilización ocurre con la fusión de dos gametos, generalmente de diferentes individuos, restaurando el estado diploide (Figura\(\PageIndex{1}\) a).
CONEXIÓN ART
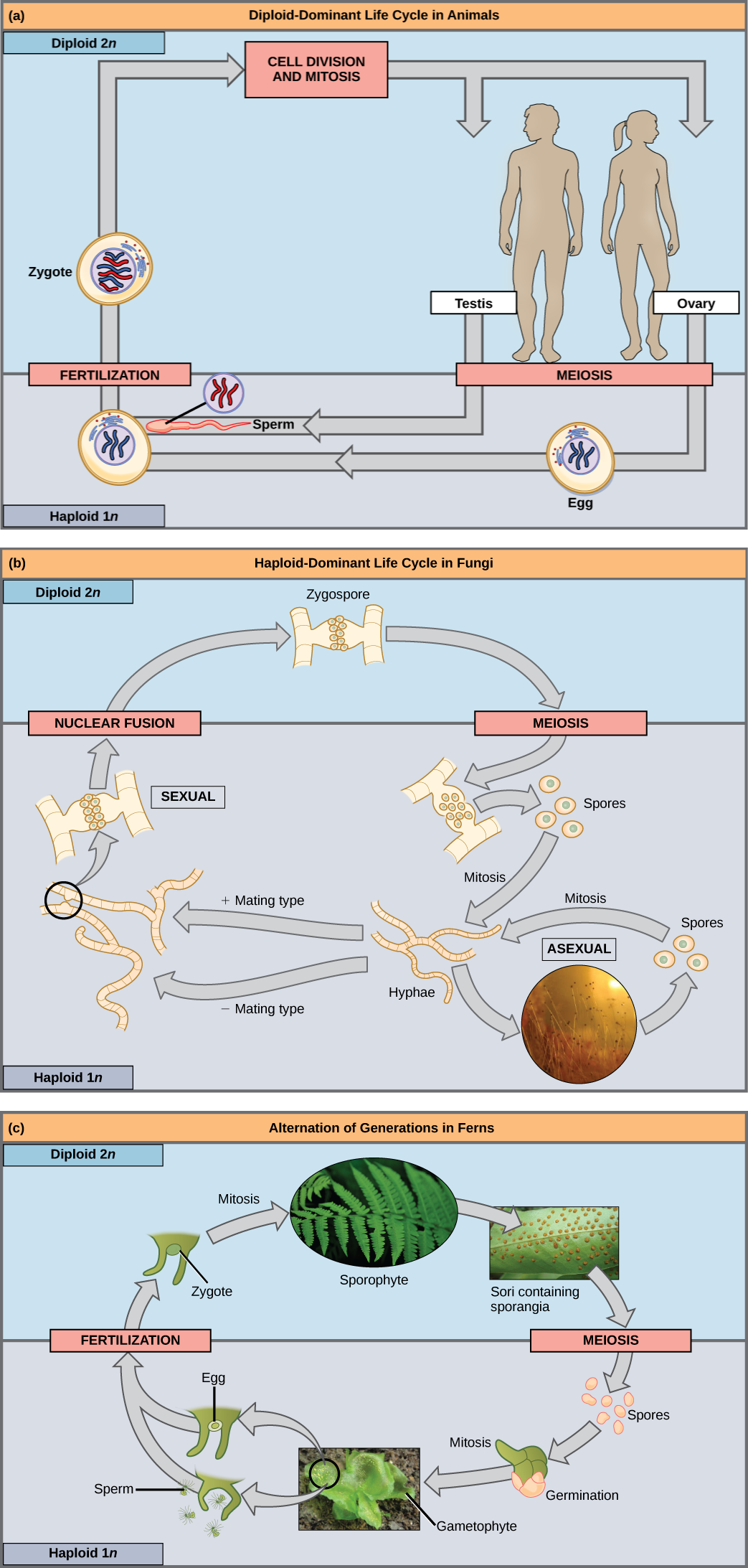
Si se produce una mutación para que un hongo ya no pueda producir un tipo de apareamiento negativo, ¿seguirá siendo capaz de reproducirse?
La mayoría de hongos y algas emplean una estrategia de ciclo de vida en la que el “cuerpo” multicelular del organismo es haploide. Durante la reproducción sexual, células haploides especializadas de dos individuos se unen para formar un cigoto diploide. El cigoto se somete inmediatamente a meiosis para formar cuatro células haploides llamadas esporas (Figura\(\PageIndex{1}\) b).
El tercer tipo de ciclo de vida, empleado por algunas algas y todas las plantas, se llama alternancia de generaciones. Estas especies tienen organismos multicelulares tanto haploides como diploides como parte de su ciclo de vida. Las plantas haploides multicelulares se llaman gametofitos porque producen gametos. La meiosis no está involucrada en la producción de gametos en este caso, ya que el organismo que produce gametos ya es haploide. La fertilización entre los gametos forma un cigoto diploide. El cigoto sufrirá muchas rondas de mitosis y dará lugar a una planta multicelular diploide llamada esporófito. Las células especializadas del esporofito sufrirán meiosis y producirán esporas haploides. Las esporas se desarrollarán en los gametofitos (Figura\(\PageIndex{1}\) c).
Resumen de la Sección
Casi todos los eucariotas se someten a reproducción sexual. La variación introducida en las células reproductivas por la meiosis parece ser una de las ventajas de la reproducción sexual que la ha hecho tan exitosa. La meiosis y la fertilización se alternan en los ciclos de vida sexual. El proceso de meiosis produce células reproductivas genéticamente únicas llamadas gametos, que tienen la mitad del número de cromosomas que la célula madre. La fertilización, la fusión de gametos haploides de dos individuos, restaura la condición diploide. Así, los organismos de reproducción sexual se alternan entre los estadios haploide y diploide. Sin embargo, las formas en que se producen las células reproductivas y el tiempo entre la meiosis y la fertilización varían mucho. Existen tres categorías principales de ciclos de vida: diploide-dominante, demostrada por la mayoría de los animales; haploide dominante, demostrada por todos los hongos y algunas algas; y alternancia de generaciones, demostrada por plantas y algunas algas.
Conexiones de arte
Figura\(\PageIndex{1}\): Si ocurre una mutación para que un hongo ya no pueda producir un tipo de apareamiento negativo, ¿seguirá siendo capaz de reproducirse?
- Contestar
-
Sí, podrá reproducirse asexualmente.
Notas al pie
- 1 Leigh Van Valen, “Una nueva ley evolutiva”, Teoría Evolutiva 1 (1973): 1—30.
Glosario
- alternancia de generaciones
- un tipo de ciclo de vida en el que se alternan las etapas diploide y haploide
- diploide-dominante
- un tipo de ciclo de vida en el que prevalece el estadio diploide multicelular
- haploide-dominante
- un tipo de ciclo de vida en el que prevalece el estadio haploide multicelular
- gametofitos
- una etapa del ciclo de vida haploide multicelular que produce gametos
- célula germinal
- una célula especializada que produce gametos, como óvulos o espermatozoides
- ciclo de vida
- la secuencia de eventos en el desarrollo de un organismo y la producción de células que producen descendencia
- meiosis
- un proceso de división nuclear que da como resultado cuatro células haploides
- esporófito
- una etapa del ciclo de vida diploide multicelular que produce esporas