
8.3: Prórrogas de las Leyes de Sucesiones
- Page ID
- 53618

Mendel estudió rasgos con un solo modo de herencia en plantas de guisante. La herencia de los rasgos que estudió siguió el patrón relativamente simple de alelos dominantes y recesivos para una sola característica. Existen varios modos importantes de herencia, descubiertos después de la obra de Mendel, que no siguen el modelo dominante y recesivo, de un solo gen.
Alternativas al dominio y la recesividad
Los experimentos de Mendel con plantas de guisante sugirieron que: 1) existen dos tipos de “unidades” o alelos para cada gen; 2) los alelos mantienen su integridad en cada generación (sin mezcla); y 3) en presencia del alelo dominante, el alelo recesivo está oculto, sin contribución al fenotipo. Por lo tanto, los alelos recesivos pueden ser “portados” y no expresados por individuos. Tales individuos heterocigóticos a veces se denominan “portadores”. Desde entonces, los estudios genéticos en otros organismos han demostrado que existe mucha más complejidad, pero que los principios fundamentales de la genética mendeliana siguen siendo ciertos. En las secciones a seguir, consideramos algunas de las extensiones del Mendelismo.
Dominio Incompleto
Los resultados de Mendel, demostrando que los rasgos son heredados como pares dominantes y recesivos, contradicen la visión en ese momento de que la descendencia exhibía una mezcla de los rasgos de sus padres. Sin embargo, el fenotipo heterocigoto ocasionalmente parece ser intermedio entre los dos progenitores. Por ejemplo, en la boca de dragón, Antirrhinum majus (Figura\(\PageIndex{1}\)), un cruce entre un progenitor homocigótico con flores blancas (C W C W) y un progenitor homocigótico con flores rojas (C R C R) producirá descendencia con rosa flores (C R C W). (Tenga en cuenta que se utilizan diferentes abreviaturas genotípicas para las extensiones mendelianas para distinguir estos patrones de dominancia simple y recesividad). Este patrón de herencia se describe como dominancia incompleta, lo que significa que uno de los alelos aparece en el fenotipo en el heterocigoto, pero no con exclusión del otro, lo que también se puede observar. El alelo de las flores rojas es incompletamente dominante sobre el alelo de las flores blancas. Sin embargo, aún se pueden predecir los resultados de un autocruzamiento heterocigoto, al igual que con los cruces mendelianos dominantes y recesivos. En este caso, la relación genotípica sería de 1 C R C R:2 C R C W:1 C W C W, y la relación fenotípica sería 1:2:1 para rojo:rosa:blanco. La base del color intermedio en el heterocigoto es simplemente que el pigmento producido por el alelo rojo (antocianina) se diluye en el heterocigoto y por lo tanto aparece rosado por el fondo blanco de los pétalos de las flores.

Codominancia
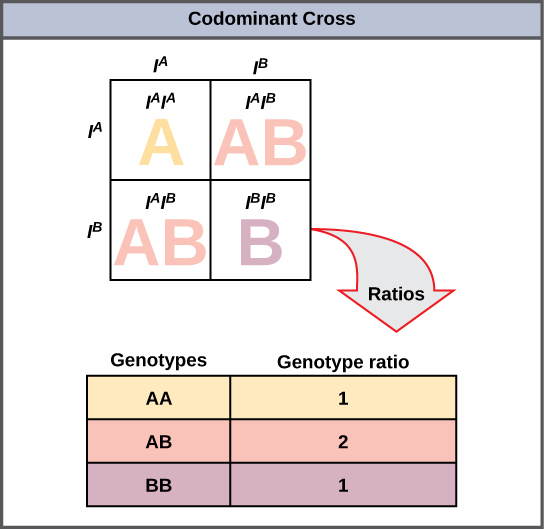
Una variación sobre la dominancia incompleta es la codominancia, en la que ambos alelos para una misma característica se expresan simultáneamente en el heterocigoto. Un ejemplo de codominancia ocurre en los grupos sanguíneos ABO de humanos. Los alelos A y B se expresan en forma de moléculas A o B presentes en la superficie de los glóbulos rojos. Los homocigotos (I A I A y I B I B) expresan el fenotipo A o B, y los heterocigotos (I A I B) expresan ambos fenotipos por igual. El individuo I A I B tiene el tipo sanguíneo AB. En un autocruce entre heterocigotos que expresan un rasgo codominante, los tres posibles genotipos de crías son fenotípicamente distintos. Sin embargo, aún se aplica la relación genotípica 1:2:1 característica de un cruce monohíbrido mendeliano (Figura\(\PageIndex{2}\)).
Alelos múltiples
Mendel implicaba que solo podían existir dos alelos, uno dominante y otro recesivo para un gen dado. Ahora sabemos que se trata de una simplificación excesiva. Aunque los humanos individuales (y todos los organismos diploides) solo pueden tener dos alelos para un gen dado, pueden existir múltiples alelos a nivel poblacional, de tal manera que se observan muchas combinaciones de dos alelos. Tenga en cuenta que cuando existen muchos alelos para un mismo gen, la convención es denotar el fenotipo o genotipo más común en la población natural como el tipo silvestre (a menudo abreviado “+”). Todos los demás fenotipos o genotipos se consideran variantes (mutantes) de esta forma típica, lo que significa que se desvían del tipo silvestre. La variante puede ser recesiva o dominante con respecto al alelo de tipo silvestre.
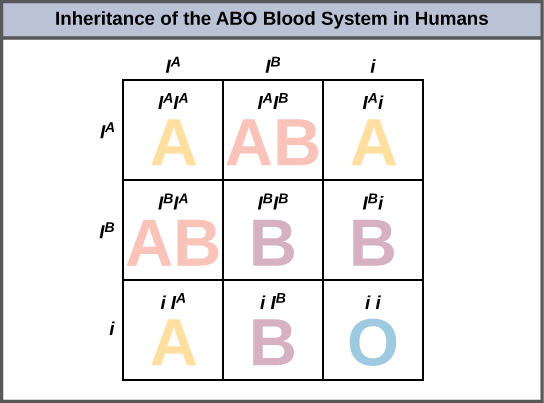
Un ejemplo de alelos múltiples es el sistema de tipo sanguíneo ABO en humanos. En este caso, hay tres alelos circulando en la población. El alelo I A codifica moléculas A en los glóbulos rojos, el alelo I B codifica moléculas B en la superficie de los glóbulos rojos y el alelo i codifica para ninguna molécula en los glóbulos rojos. En este caso, los alelos I A y I B son codominantes entre sí y ambos son dominantes sobre el alelo i. Si bien hay tres alelos presentes en una población, cada individuo sólo obtiene dos de los alelos de sus padres. Esto produce los genotipos y fenotipos mostrados en la Figura\(\PageIndex{3}\). Observe que en lugar de tres genotipos, hay seis genotipos diferentes cuando hay tres alelos. El número de fenotipos posibles depende de las relaciones de dominancia entre los tres alelos.
EVOLUCIÓN EN ACCIÓN: Alelos Múltiples confieren Resistencia a Fármacos en el Parásito
La malaria es una enfermedad parasitaria en humanos que se transmite por mosquitos hembra infectados, incluyendo Anopheles gambiae, y se caracteriza por fiebre cíclica alta, escalofríos, síntomas similares a la gripe y anemia severa. Plasmodium falciparum y P. vivax son los agentes causantes más comunes de la malaria, y P. falciparum es el más mortal. Cuando se trata rápida y correctamente, la malaria por P. falciparum tiene una tasa de mortalidad de 0.1 por ciento. Sin embargo, en algunas partes del mundo, el parásito ha evolucionado resistencia a los tratamientos de malaria de uso común, por lo que los tratamientos palúdicos más efectivos pueden variar según la región geográfica.
En el sudeste asiático, África y Sudamérica, P. falciparum ha desarrollado resistencia a los fármacos antipalúdicos cloroquina, mefloquina y sulfadoxina-pirimetamina. P. falciparum, que es haploide durante la etapa de vida en la que es infeccioso para los humanos, ha evolucionado múltiples alelos mutantes farmacorresistentes del gen dhps. Diferentes grados de resistencia a la sulfadoxina están asociados con cada uno de estos alelos. Al ser haploide, P. falciparum solo necesita un alelo farmacorresistente para expresar este rasgo.
En el sudeste asiático, diferentes alelos resistentes a sulfadoxina del gen dhps se localizan en diferentes regiones geográficas. Este es un fenómeno evolutivo común que se produce porque los mutantes farmacorresistentes surgen en una población y se cruzan con otros aislados de P. falciparum en las proximidades. Los parásitos resistentes a la sulfadoxina causan considerables dificultades humanas en regiones en las que este medicamento es ampliamente utilizado como remedio contra la malaria de venta libre. Como es común con patógenos que se multiplican a grandes números dentro de un ciclo de infección, P. falciparum evoluciona relativamente rápido (más de una década más o menos) en respuesta a la presión selectiva de los fármacos antipalúdicos de uso común. Por esta razón, los científicos deben trabajar constantemente para desarrollar nuevos medicamentos o combinaciones de medicamentos para combatir la carga mundial de malaria. 1
Rasgos vinculados al sexo
En humanos, así como en muchos otros animales y algunas plantas, el sexo del individuo está determinado por cromosomas sexuales, un par de cromosomas no homólogos. Hasta ahora, solo hemos considerado patrones de herencia entre cromosomas no sexuales, o autosomas. Además de 22 pares homólogos de autosomas, las hembras humanas tienen un par homólogo de cromosomas X, mientras que los machos humanos tienen un par de cromosomas XY. Aunque el cromosoma Y contiene una pequeña región de similitud con el cromosoma X para que puedan emparejarse durante la meiosis, el cromosoma Y es mucho más corto y contiene menos genes. Cuando un gen que se examina está presente en el cromosoma X, pero no en el cromosoma Y, se encuentra ligado al X.
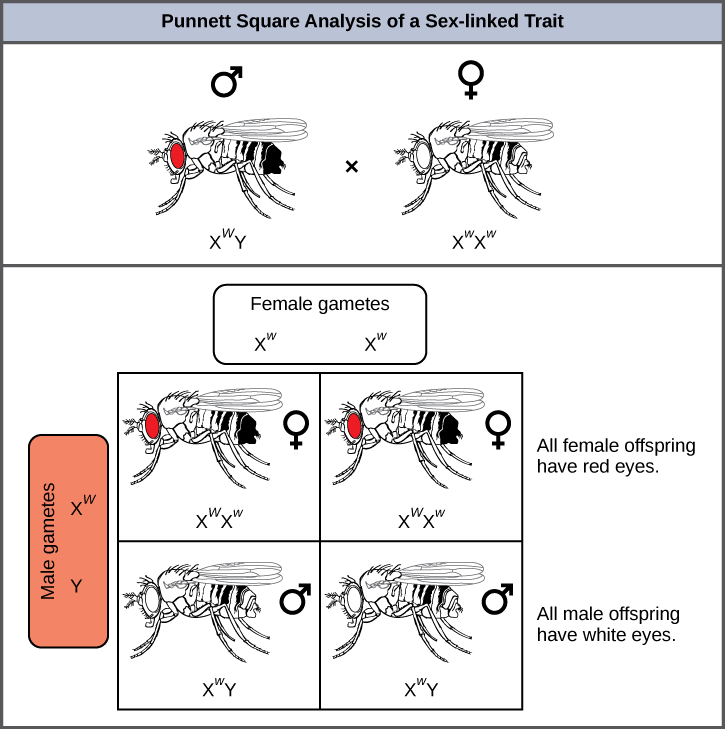
El color de ojos en Drosophila, la mosca común de la fruta, fue el primer rasgo ligado al cromosoma X que se identificó. Thomas Hunt Morgan mapeó este rasgo al cromosoma X en 1910. Al igual que los humanos, los machos de Drosophila tienen un par de cromosomas XY y las hembras son XX. En las moscas el color de ojos de tipo silvestre es rojo (X W) y es dominante al color de ojos blanco (X w) (Figura\(\PageIndex{4}\)). Debido a la ubicación del gen del color de los ojos, los cruces recíprocos no producen las mismas proporciones de descendencia. Se dice que los machos son hemicigóticos, ya que tienen un solo alelo para cualquier característica ligada a X. La hemicigosidad hace que las descripciones de dominancia y recesividad sean irrelevantes para los machos XY. Los machos de Drosophila carecen del gen blanco en el cromosoma Y; es decir, su genotipo solo puede ser X W Y o X w Y. En contraste, las hembras tienen dos copias alélicas de este gen y pueden ser X W X W, X W X w, o X w X w.

En un cruce ligado al X, los genotipos de las crías F 1 y F 2 dependen de si el rasgo recesivo fue expresado por el macho o la hembra en la generación P. Con respecto al color de ojos de Drosophila, cuando el macho P expresa el fenotipo de ojo blanco y la hembra tiene ojos rojos homocigóticos, todos los miembros de la generación F 1 presentan ojos rojos (Figura\(\PageIndex{5}\)). Las hembras F 1 son heterocigóticas (X W X w), y los machos son todos X W Y, habiendo recibido su cromosoma X de la hembra P dominante homocigótica y su cromosoma Y del macho P. Un cruce posterior entre la hembra X W X w y el macho X W Y produciría solo hembras de ojos rojos (con X W X W o X W X w) y machos de ojos rojos y blancos (con genotipos X W Y o X w Y). Ahora, consideremos un cruce entre una hembra homocigótica de ojos blancos y un macho con ojos rojos. La generación F 1 exhibiría solo hembras heterocigóticas de ojos rojos (X W X w) y solo machos de ojos blancos (X w Y). La mitad de las hembras F 2 serían de ojos rojos (X W X w) y la mitad serían de ojos blancos (X w X w). De igual manera, la mitad de los machos F 2 serían de ojos rojos (X W Y) y la mitad serían de ojos blancos (X w Y).
CONEXIÓN ARTE
¿Qué proporción de descendencia resultaría de un cruce entre un macho de ojos blancos y una hembra heterocigoto para el color de ojos rojos?
Los descubrimientos en genética de la mosca de la fruta se pueden aplicar a la genética humana. Cuando una madre femenina es homocigótica para un rasgo recesivo ligado al X, pasará el rasgo al 100 por ciento de su descendencia masculina, porque los machos recibirán el cromosoma Y del progenitor masculino. En humanos, los alelos para ciertas afecciones (algunos daltonismo, hemofilia y distrofia muscular) están ligados al cromosoma X. Se dice que las hembras heterocigóticas para estas enfermedades son portadoras y pueden no exhibir ningún efecto fenotípico. Estas hembras pasarán la enfermedad a la mitad de sus hijos y pasarán el estado de portadora a la mitad de sus hijas; por lo tanto, los rasgos ligados al X aparecen con mayor frecuencia en hombres que en mujeres.
En algunos grupos de organismos con cromosomas sexuales, el sexo con los cromosomas sexuales no homólogos es el femenino más que el masculino. Este es el caso de todas las aves. En este caso, los rasgos vinculados al sexo tendrán más probabilidades de aparecer en la hembra, en la que son hemicigóticos.
CONCEPT EN ACCIÓN
Mira este video para conocer más sobre los rasgos vinculados al sexo.
Genes Vinculados Violan la Ley del Surtido Independiente
Aunque todas las características de la planta de guisante de Mendel se comportaron de acuerdo con la ley del surtido independiente, ahora sabemos que algunas combinaciones de alelos no se heredan independientemente entre sí. Los genes que se localizan en cromosomas separados y no homólogos siempre se ordenarán de forma independiente. Sin embargo, cada cromosoma contiene cientos o miles de genes, organizados linealmente en cromosomas como perlas en una cuerda. La segregación de alelos en gametos puede verse influenciada por el ligamiento, en el que los genes que se localizan físicamente cerca unos de otros en el mismo cromosoma tienen más probabilidades de ser heredados como pareja. Sin embargo, debido al proceso de recombinación, o “cruce”, es posible que dos genes en un mismo cromosoma se comporten de forma independiente, o como si no estuvieran vinculados. Para entender esto, consideremos las bases biológicas de la unión génica y la recombinación.
Los cromosomas homólogos poseen los mismos genes en el mismo orden, aunque los alelos específicos del gen pueden ser diferentes en cada uno de los dos cromosomas. Recordemos que durante la interfase y profase I de la meiosis, los cromosomas homólogos primero se replican y luego la sinapsis, con genes similares en los homólogos alineándose entre sí. En esta etapa, segmentos de cromosomas homólogos intercambian segmentos lineales de material genético (Figura\(\PageIndex{6}\)). Este proceso se llama recombinación, o cruce, y es un proceso genético común. Debido a que los genes están alineados durante la recombinación, el orden de los genes no se altera. En cambio, el resultado de la recombinación es que los alelos maternos y paternos se combinan en el mismo cromosoma. A lo largo de un cromosoma dado, pueden ocurrir varios eventos de recombinación, lo que provoca un extenso barajado de alelos.
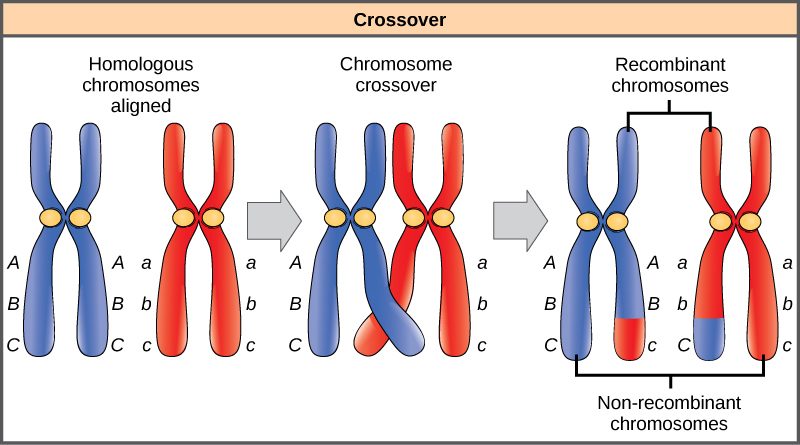
Cuando dos genes se localizan en un mismo cromosoma, se consideran vinculados, y sus alelos tienden a transmitirse a través de la meiosis juntos. Para ejemplificar esto, imagine un cruce dihíbrido que involucre el color de la flor y la altura de la planta en el que los genes están uno al lado del otro en el cromosoma. Si un cromosoma homólogo tiene alelos para plantas altas y flores rojas, y el otro cromosoma tiene genes para plantas cortas y flores amarillas, entonces cuando se forman los gametos, los alelos altos y rojos tenderán a ir juntos en un gameto y los alelos cortos y amarillos irán a otros gametos. Estos se denominan genotipos parentales porque han sido heredados intactos de los padres del individuo productor de gametos. Pero a diferencia de si los genes estuvieran en diferentes cromosomas, no habrá gametos con alelos altos y amarillos ni gametos con alelos cortos y rojos. Si creas un cuadrado Punnett con estos gametos, verás que no se aplicaría la predicción mendeliana clásica de un resultado 9:3:3:1 de un cruce dihíbrido. A medida que aumenta la distancia entre dos genes, aumenta la probabilidad de uno o más cruces entre ellos y los genes se comportan más como si estuvieran en cromosomas separados. Los genetistas han utilizado la proporción de gametos recombinantes (a los que no les gustan los padres) como medida de lo lejos que están los genes en un cromosoma. Usando esta información, han construido mapas de ligamiento de genes en cromosomas para organismos bien estudiados, incluidos los humanos.
La publicación seminal de Mendel no hace mención de vinculación, y muchos investigadores han cuestionado si encontró vinculación pero optaron por no publicar esos cruces por preocupación de que invalidarían su postulado de surtido independiente. El guisante de jardín tiene siete cromosomas, y algunos han sugerido que su elección de siete características no fue una coincidencia. Sin embargo, aunque los genes que examinó no estuvieran localizados en cromosomas separados, es posible que simplemente no observara el enlace debido a los extensos efectos de barajado de la recombinación.
Epistasis
Los estudios de Mendel en plantas de guisante implicaban que la suma del fenotipo de un individuo estaba controlada por genes (o como él los llamaba, factores unitarios), de tal manera que cada característica estaba clara y completamente controlada por un solo gen. De hecho, las características observables individuales casi siempre están bajo la influencia de múltiples genes (cada uno con dos o más alelos) que actúan al unísono. Por ejemplo, al menos ocho genes contribuyen al color de los ojos en los humanos.
CONCEPT EN ACCIÓN

El color de ojos en humanos está determinado por múltiples alelos. Usa la Calculadora de Color de Ojos para predecir el color de ojos de los niños a partir del color de ojos de los
En algunos casos, varios genes pueden contribuir a aspectos de un fenotipo común sin que sus productos génicos interactúen directamente. En el caso del desarrollo de órganos, por ejemplo, los genes pueden expresarse secuencialmente, añadiendo cada gen a la complejidad y especificidad del órgano. Los genes pueden funcionar de manera complementaria o sinérgica, de tal manera que dos o más genes expresados simultáneamente afectan a un fenotipo. Un ejemplo aparente de esto ocurre con el color de la piel humana, que parece implicar la acción de al menos tres (y probablemente más) genes. Los casos en los que la herencia por una característica como el color de la piel o la altura humana dependen de los efectos combinados de numerosos genes se denominan herencia poligénica.
Los genes también pueden oponerse entre sí, con un gen suprimiendo la expresión de otro. En epistasis, la interacción entre genes es antagónica, de tal manera que un gen enmascara o interfiere con la expresión de otro. “Epistasis” es una palabra compuesta de raíces griegas que significa “de pie sobre”. Se dice que los alelos que se están enmascarando o silenciando son hipostáticos a los alelos epistáticos que están haciendo el enmascaramiento. A menudo, la base bioquímica de la epistasis es una vía génica en la que la expresión de un gen depende de la función de un gen que lo precede o sigue en la ruta.
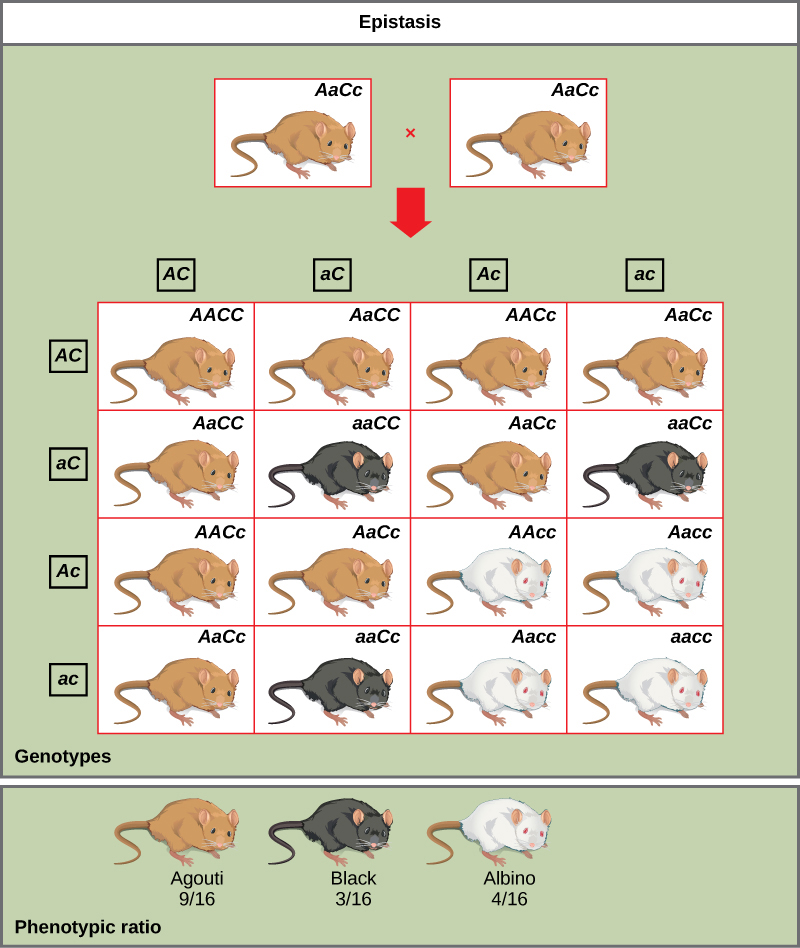
Un ejemplo de epistasis es la pigmentación en ratones. El color del pelaje de tipo silvestre, aguti (AA) es dominante al pelaje de color sólido (aa). Sin embargo, un gen C separado, cuando está presente como el homocigoto recesivo (cc), niega cualquier expresión de pigmento del gen A y da como resultado un ratón albino (Figura\(\PageIndex{7}\)). Por lo tanto, los genotipos AacC, Aacc y aacc producen el mismo fenotipo albino. Un cruce entre heterocigotos para ambos genes (AacC x AACc) generaría descendencia con una relación fenotípica de 9 aguti:3 negro:4 albino (Figura\(\PageIndex{7}\)). En este caso, el gen C es epistático al gen A.
Resumen de la Sección
Los alelos no siempre se comportan en patrones dominantes y recesivos. La dominancia incompleta describe situaciones en las que el heterocigoto exhibe un fenotipo que es intermedio entre los fenotipos homocigóticos. La codominancia describe la expresión simultánea de ambos alelos en el heterocigoto. Aunque los organismos diploides solo pueden tener dos alelos para un gen dado, es común que más de dos alelos exista un gen en una población. En los humanos, como en muchos animales y algunas plantas, las hembras tienen dos cromosomas X y los machos tienen un cromosoma X y un cromosoma Y. Se dice que los genes que están presentes en el cromosoma X pero no en el cromosoma Y están ligados a X, de tal manera que los machos solo heredan un alelo para el gen, y las hembras heredan dos.
Según la ley de Mendel de surtido independiente, los genes se clasifican independientemente entre sí en gametos durante la meiosis. Esto ocurre porque los cromosomas, en los que residen los genes, se asumen independientemente durante la meiosis y los cruces hacen que la mayoría de los genes en los mismos cromosomas también se comporten de manera independiente. Cuando los genes se localizan muy cerca en el mismo cromosoma, sus alelos tienden a heredarse juntos. Esto da como resultado proporciones de descendencia que violan la ley de surtido independiente de Mendel. Sin embargo, la recombinación sirve para intercambiar material genético en cromosomas homólogos de tal manera que los alelos maternos y paternos pueden recombinarse en el mismo cromosoma. Es por ello que los alelos en un cromosoma dado no siempre se heredan juntos. La recombinación es un evento aleatorio que ocurre en cualquier parte de un cromosoma. Por lo tanto, es probable que los genes que están muy separados en el mismo cromosoma sigan asestando independientemente debido a eventos de recombinación que ocurrieron en el espacio cromosómico intermedio.
Se estén clasificando o no de forma independiente, los genes pueden interactuar a nivel de productos génicos, de tal manera que la expresión de un alelo para un gen enmascara o modifica la expresión de un alelo para un gen diferente. A esto se le llama epistasis.
Conexiones de arte
Figura\(\PageIndex{5}\): ¿Qué relación de descendencia resultaría de un cruce entre un macho de ojos blancos y una hembra heterocigoto para el color de ojos rojos?
- Contestar
-
La mitad de la descendencia femenina sería heterocigótica (X W X w) con ojos rojos, y la mitad sería homocigótica recesiva (X w X w) con ojos blancos. La mitad de la descendencia masculina sería dominante hemicigótica (X W Y) con ojos rojos, y la mitad sería hemicigótica recesiva (X w Y) con ojos blancos.
Notas al pie
- 1 Sumiti Vinayak et al., “Origen y Evolución de Plasmodium falciparum Resistente a Sulfadoxina”, PLoS Patógenos 6 (2010): e1000830.
Glosario
- codominancia
- en un heterocigoto, expresión completa y simultánea de ambos alelos para una misma característica
- epistasis
- una interacción entre genes de tal manera que un gen enmascara o interfiere con la expresión de otro
- hemicigótico
- la presencia de un solo alelo para una característica, como en el ligamiento X; la hemicigosidad hace irrelevantes las descripciones de dominancia y recesividad
- dominio incompleto
- en un heterocigoto, expresión de dos alelos contrastantes de manera que el individuo muestre un fenotipo intermedio
- vinculación
- un fenómeno en el que los alelos que se encuentran muy cerca unos de otros en el mismo cromosoma tienen más probabilidades de heredarse juntos
- recombinación
- el proceso durante la meiosis en el que los cromosomas homólogos intercambian segmentos lineales de material genético, aumentando drásticamente la variación genética en la descendencia y separando los genes enlazados
- tipo salvaje
- el genotipo o fenotipo más frecuente para una característica dada que se encuentra en una población
- Vinculado a X
- un gen presente en el cromosoma X, pero no en el cromosoma Y