
3.6: Lípidos y Membranas
- Page ID
- 53647

Los lípidos son una amplia clase de moléculas que comparten la característica de que tienen al menos una porción de ellas que es hidrófoba. La clase de moléculas incluye grasas, aceites (y sus ácidos grasos sustituyentes), esteroides, vitaminas liposolubles, prostaglandinas, glicerofosfolípidos y esfingolípidos. Curiosamente, cada uno de estos puede derivarse de acetil-CoA Podría decirse que los lípidos más importantes en nuestras células son los ácidos grasos, porque son componentes de todos los demás lípidos, excepto algunos de los esteroides y vitaminas liposolubles. Constituidos por un grupo carboxilo unido a una larga cola alifática, los ácidos grasos se describen como saturados (sin dobles enlaces) o insaturados (uno o más dobles enlaces). Los ácidos grasos con más de un doble enlace se describen como poliinsaturados. Al aumentar la cantidad de ácidos grasos insaturados (y la cantidad de insaturación en un ácido graso dado) en una grasa disminuye su temperatura de fusión. Esto también es un factor en la fluidez de la membrana. Si la temperatura de fusión de una grasa disminuye suficientemente para que sea un líquido a temperatura ambiente, se le conoce como un aceite. Cabe destacar que organismos como los peces, que viven en ambientes fríos, tienen grasas con más insaturación. Es por ello que el aceite de pescado es una rica fuente de ácidos grasos poliinsaturados.
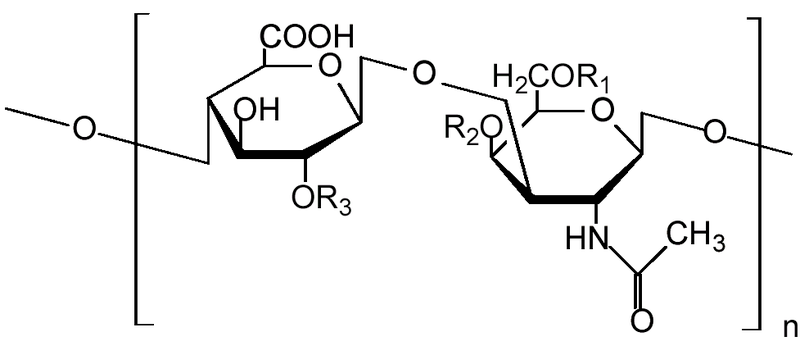
Figura\(\PageIndex{1}\): Unidad monomérica de sulfato de condroitina. Estructura química de una unidad en una cadena de sulfato de condroitina.
Ácidos grasos
Podría decirse que los lípidos más importantes en nuestras células son los ácidos grasos, ya que son componentes de todos los demás lípidos, excepto algunos de los esteroides y vitaminas liposolubles. Constituidos por un grupo carboxilo unido a una larga cola alifática, los ácidos grasos se describen como saturados (sin dobles enlaces) o insaturados (uno o más dobles enlaces). Los ácidos grasos con más de un doble enlace se describen como poliinsaturados. Al aumentar la cantidad de ácidos grasos insaturados (y la cantidad de insaturación en un ácido graso dado) en una grasa disminuye su temperatura de fusión. Esto también es un factor en la fluidez de la membrana. Si la temperatura de fusión de una grasa disminuye suficientemente para que sea un líquido a temperatura ambiente, se le conoce como un aceite. Cabe destacar que organismos como los peces, que viven en ambientes fríos, tienen grasas con más insaturación. Es por ello que el aceite de pescado es una rica fuente de ácidos grasos poliinsaturados
Ácido Hialurónico
Al melodía de “Rudolph el reno de nariz roja”
Grabado por David Simmons
Letras por Kevin Ahern
Ácido Hialurónico
Actuando casi mágicamente
Colocado justo debajo de la rótula
Lubricar los escombros
Mejor que el reemplazo articular
Simple como 1-2-3
Si puede detener el dolor
Usted conseguirás mantener tu rodilla
Cuando el dolor se está poniendo mal
Intenta no estar triste
Solo sal a platicar
Con tu médico ortopédico
Pídele que use la aguja
Para no hacerlo sería un delito
Ácido hialurónico
Trabajando donde el sol no brilla
Mesa\(\PageIndex{1}\): De arriba a abajo - Vitaminas E, K y A
| Número de átomos de carbono | Insaturación | Fórmula |
Punto de fusión |
|
|---|---|---|---|---|
| Palmitoleico | 16 | \( 16: 1-\Delta^{9}\) | \(\mathrm{CH}_{3}\left(\mathrm{CH}_{2}\right)_{5} \mathrm{CH}=\mathrm{CH}\left(\mathrm{CH}_{2}\right)_{7} \mathrm{CO}_{2} \mathrm{H}\) | -0.5 |
| Oleico | 18 | \( 18: 1-\Delta^{9}\) | \(\mathrm{CH}_{3}\left(\mathrm{CH}_{2}\right)_{7} \mathrm{CH}=\mathrm{CH}\left(\mathrm{CH}_{2}\right)_{7} \mathrm{CO}_{2} \mathrm{H}\) | 16 |
| Linoleico | 18 | \( 18: 2-\Delta^{9,12}\) | \(\mathrm{CH}_{3}\left(\mathrm{CH}_{2}\right)_{4} \mathrm{CH}=\mathrm{CH}\left(\mathrm{CH}_{2}\right) \mathrm{CH}=\mathrm{CH}\left(\mathrm{CH}_{2}\right)_{7} \mathrm{CO}_{2} \mathrm{H}\) | -5 |
| Linolénico | 18 | \( 18: 3-\Delta^{9,12,15}\) | \(\mathrm{CH}_{3}\left(\mathrm{CH}_{2} \mathrm{CH}=\mathrm{CH}\right)_{3}\left(\mathrm{CH}_{2}\right)_{7} \mathrm{CO}_{2} \mathrm{H}\) | -11 |
| Araquidónico | 20 | \( 20: 4-\Delta^{5,8,11,14}\) | \(\mathrm{CH}_{3}\left(\mathrm{CH}_{2}\right)_{4} \mathrm{CH}=\mathrm{CH}\left(\mathrm{CH}_{2}\right)_{4}\left(\mathrm{CH}_{2}\right)_{2} \mathrm{CO}_{2} \mathrm{H}\) | -50 |
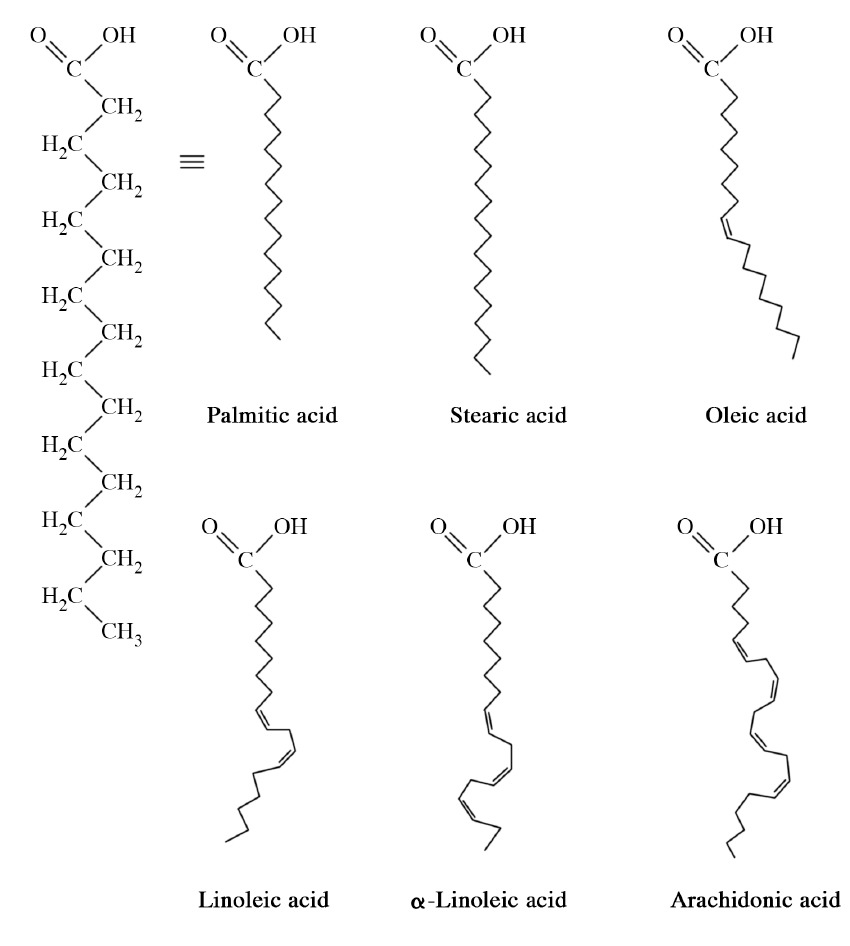
Bioquímicamente, los dobles enlaces que se encuentran en los ácidos grasos se encuentran predominantemente en la configuración cis. Las llamadas grasas trans surgen como un subproducto químico de la hidrogenación parcial del aceite vegetal (pequeñas cantidades de grasas trans también ocurren naturalmente). En humanos, el consumo de grasas trans eleva los niveles de lipoproteínas de baja densidad (LDL) y disminuye los niveles de lipoproteínas de alta densidad (HDL). Se cree que cada uno contribuye al riesgo de desarrollar enfermedad arterial coronaria. Los ácidos grasos más comunes en nuestro organismo incluyen palmitato, estearato, oleato, linolenato, linoleato y araquidonato. Los ácidos grasos están numerados por dos esquemas completamente diferentes. El esquema de numeración delta tiene el grupo carboxilo como #1, mientras que el esquema de números omega comienza en el otro extremo del ácido graso con el grupo metilo como #1. Los ácidos grasos se describen como esenciales si deben estar en la dieta (no pueden ser sintetizados por el organismo). Los animales, incluidos los humanos, no pueden sintetizar ácidos grasos con dobles enlaces más allá de la posición delta 9, por lo que los ácidos linoleico y linolénico se consideran esenciales en estos organismos. En las células animales, las grasas son las formas primarias de almacenamiento de energía. También se les conoce como triacilgliceroles, ya que consisten en una molécula de glicerol esterificada a tres ácidos grasos. Las grasas se sintetizan reemplazando el fosfato en el ácido fosfatídico por un ácido graso. Las grasas se almacenan en el cuerpo en células especializadas conocidas como adipocitos. Enzimas conocidas
ya que las lipasas liberan ácidos grasos de las grasas por reacciones de hidrólisis. De las diversas lipasas que actúan sobre la grasa, la que actúa primero, la triacilglicerol lipasa, está regulada hormonalmente.
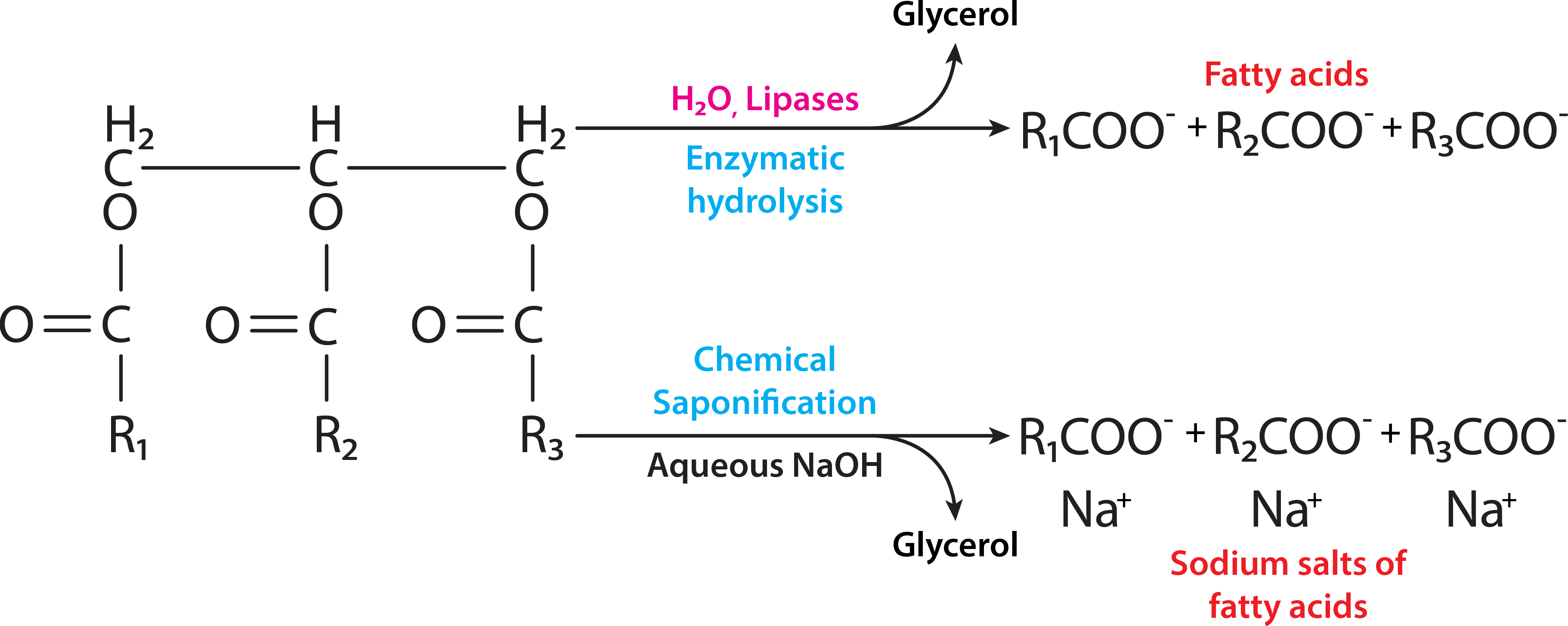
Figura\(\PageIndex{3}\): Hidrólisis de grasa
Lípidos de membrana
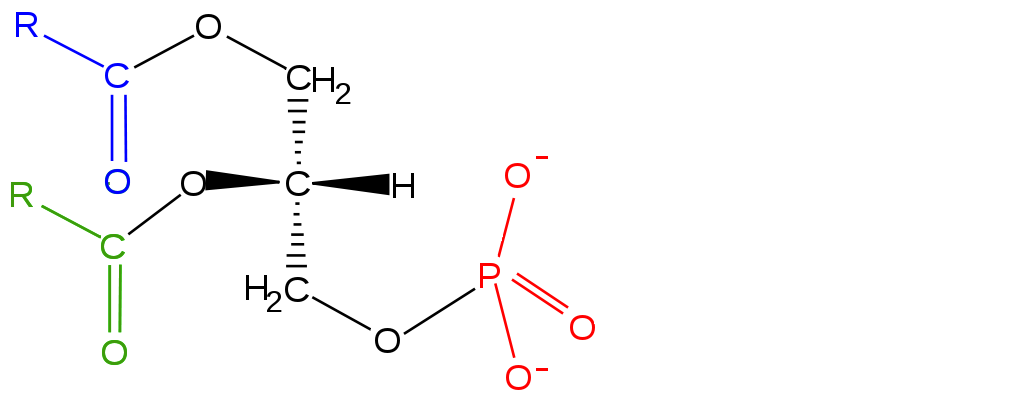
Los lípidos predominantes que se encuentran en las membranas son glicerofosfolípidos (fosfoglicéridos) y esfingolípidos. Los primeros están relacionados con las grasas estructuralmente ya que ambas se derivan del ácido fosfatídico. El ácido fosfatídico es un glicerofosfolípido simple que generalmente se convierte en compuestos de fosfatidilo. Estos se hacen esterificando diversos grupos, como etanolamina, serina, colina, inositol, y otros al fosfato. Todos estos compuestos forman bicapas lipídicas en solución acuosa, debido a la naturaleza anfifílica de su estructura. Aunque estructuralmente similares a los glicerofosfolípidos, los esfingolípidos se sintetizan completamente independientemente de ellos, comenzando con el ácido palmítico y el aminoácido serina. La figura\(\PageIndex{5}\) muestra la estructura de varios esfingolípidos.
Al igual que los glicerofosfolípidos, los esfingolípidos son anfifílicos, pero a diferencia de ellos, pueden tener carbohidratos simples (en cerebrosidos) o complejos (en gangliósidos) unidos en un extremo. La mayoría de los esfingolípidos, excepto la esfingomielina, no contienen fosfato. Los esteroides, como el colesterol, también se encuentran en las membranas. El colesterol, en particular, puede desempeñar un papel importante en la fluidez de la membrana.
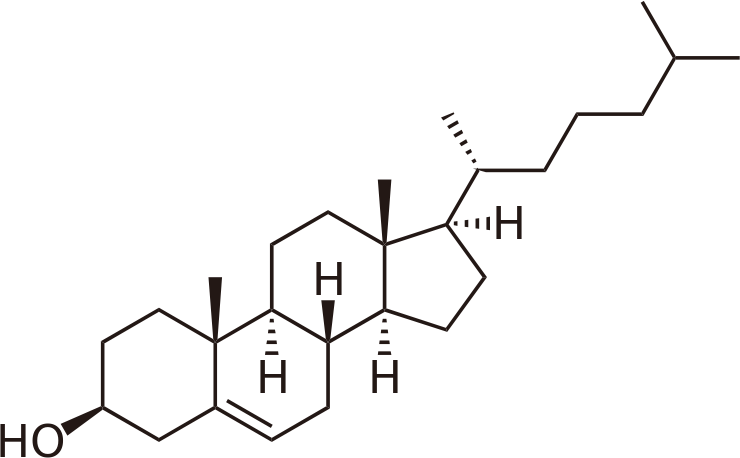
Figura\(\PageIndex{5}\): Colesterol
Las membranas se pueden pensar en un ser más “congelado” o más “fluido”. La fluidez es importante para las membranas celulares. Cuando se calientan, las membranas pasan de un carácter más “congelado” al de uno más “fluido” a medida que aumenta la temperatura. El punto medio de esta transición, referido como la Tm, está influenciado por la composición de ácidos grasos de los compuestos de bicapa lipídica. Los ácidos grasos más largos y saturados favorecerán valores de Tm más altos, mientras que la insaturación y los ácidos grasos cortos favorecerán valores de Tm menores Curiosamente, el colesterol no cambia el valor de Tm, sino que amplía el rango de transición entre las
formas congeladas y líquidas de la membrana.
Bicapas lipídicas
La membrana alrededor de las células contiene muchos componentes, incluyendo colesterol, proteínas, glicolípidos, glicerofosfolípidos y esfingolípidos. Los dos últimos de estos formarán, en el agua, lo que se denomina bicapa lipídica, que sirve como límite para la célula que es en gran parte impermeable al movimiento de la mayoría de los materiales a través de ella.
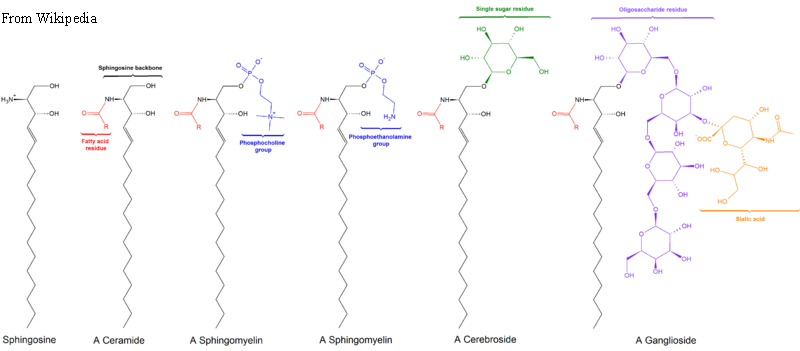
Figura\(\PageIndex{6}\): Las estructuras de varios esfingolípidos
Con las notables excepciones del agua, el dióxido de carbono, el monóxido de carbono y el oxígeno, la mayoría de los compuestos polares/iónicos requieren proteínas de transporte para ayudarlos a navegar eficientemente a través de la bicapa. El movimiento ordenado de estos compuestos es crítico para que la célula pueda 1) obtener alimentos para obtener energía; 2) exportar materiales; 3) mantener el equilibrio osmótico; 4) crear gradientes para el transporte secundario; 5) proporcionar fuerza electromotriz para la señalización nerviosa; y 6) almacenar energía en gradientes electroquímicos para ATP producción (fosforilación oxidativa o fotosíntesis). En algunos casos, se requiere energía para mover las sustancias (transporte activo). En otros casos, no se requiere energía externa y se mueven por difusión a través de canales celulares específicos.
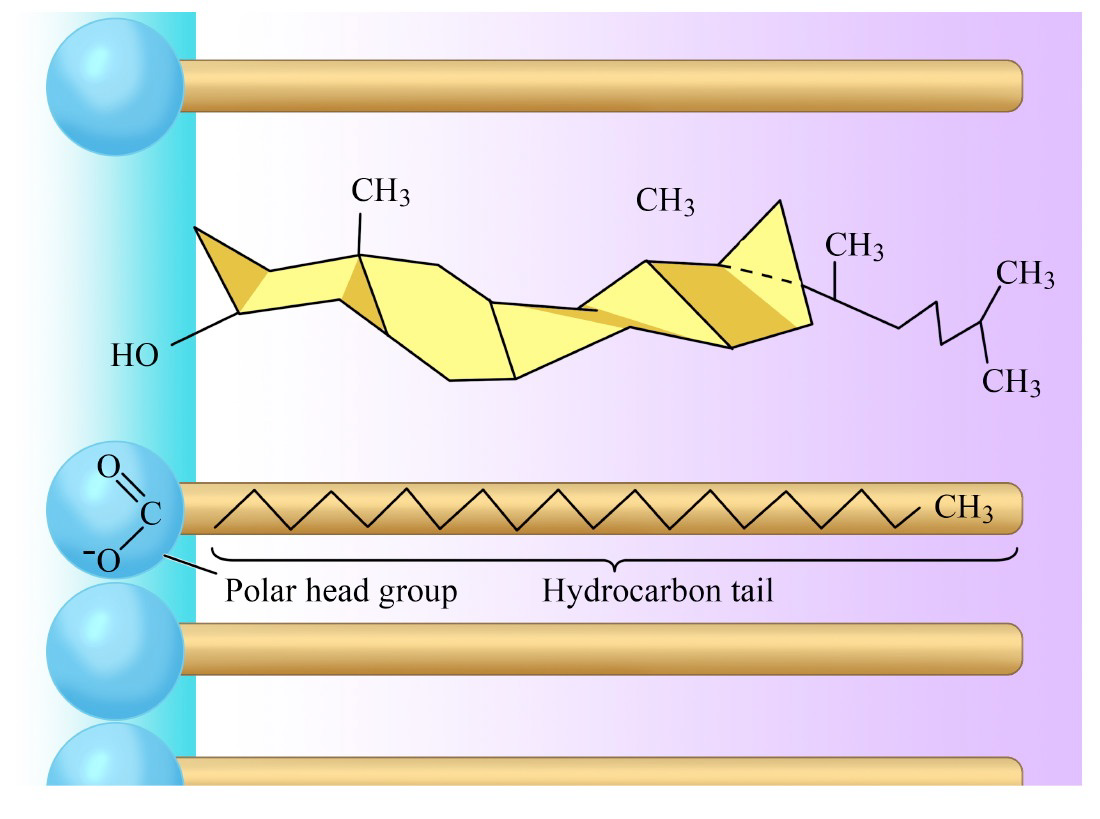
Figura\(\PageIndex{7}\): Colesterol en una bicapa lipídica
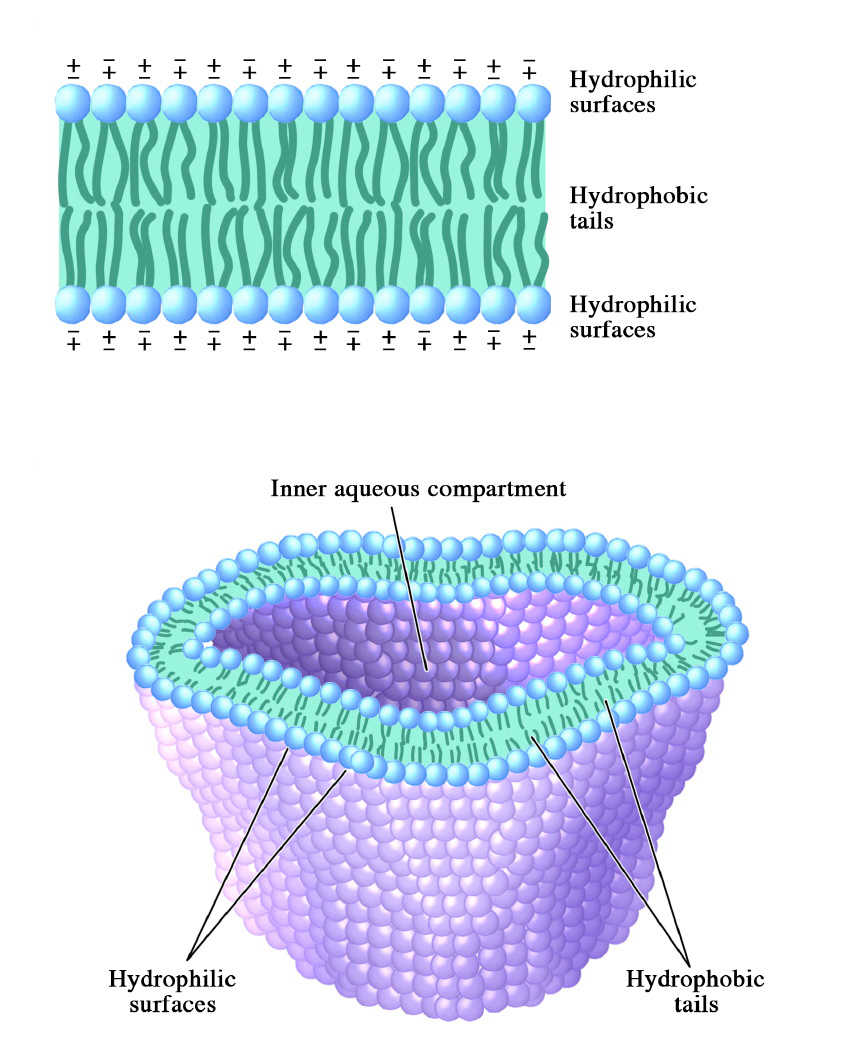
La capacidad espontánea de estos compuestos para formar bicapas lipídicas se explota en la formación de estructuras membranosas artificiales llamadas liposomas. Los liposomas tienen algunos usos para entregar su contenido a las células a través de la fusión de membrana.
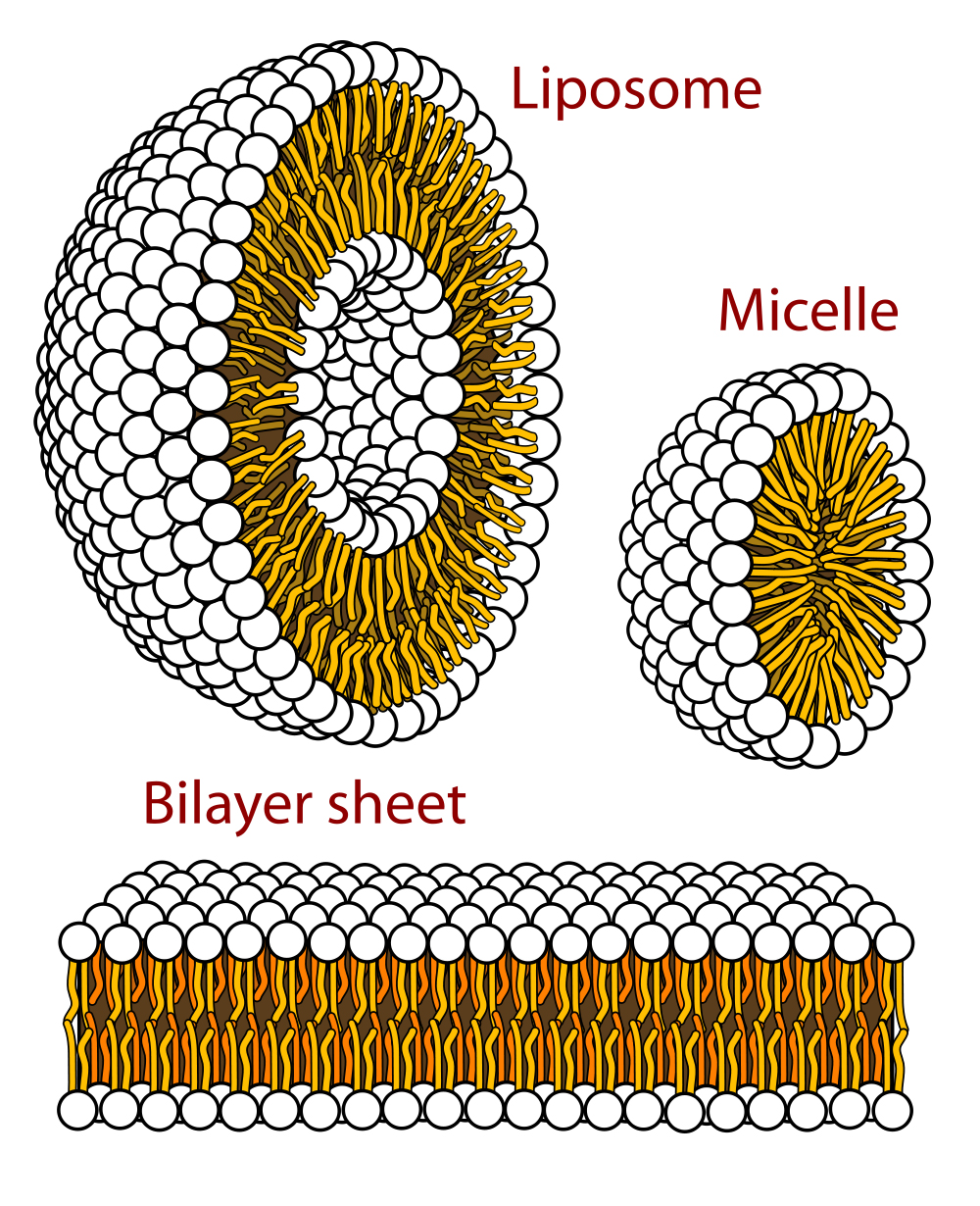
Figura\(\PageIndex{9}\): Liposomas
Proteínas de membrana
Otros componentes significativos de las membranas celulares incluyen las proteínas. Podemos ponerlos en varias categorías. Las proteínas integrales de membrana están incrustadas en la membrana y se proyectan a través de ambos lados de la bicapa lipídica. Las proteínas de membrana periférica están incrustadas o estrechamente asociadas con parte de la bicapa, pero no se proyectan completamente a través de ambos lados. Las proteínas de membrana asociadas se encuentran cerca de las membranas, pero pueden no estar incrustadas en ellas. Su asociación puede surgir como resultado de la interacción con otras proteínas o moléculas en la bicapa lipídica. Las proteínas de membrana ancladas no están incrustadas en la bicapa lipídica, sino que están unidas a una molécula (típicamente un ácido graso) que está incrustada en la membrana.
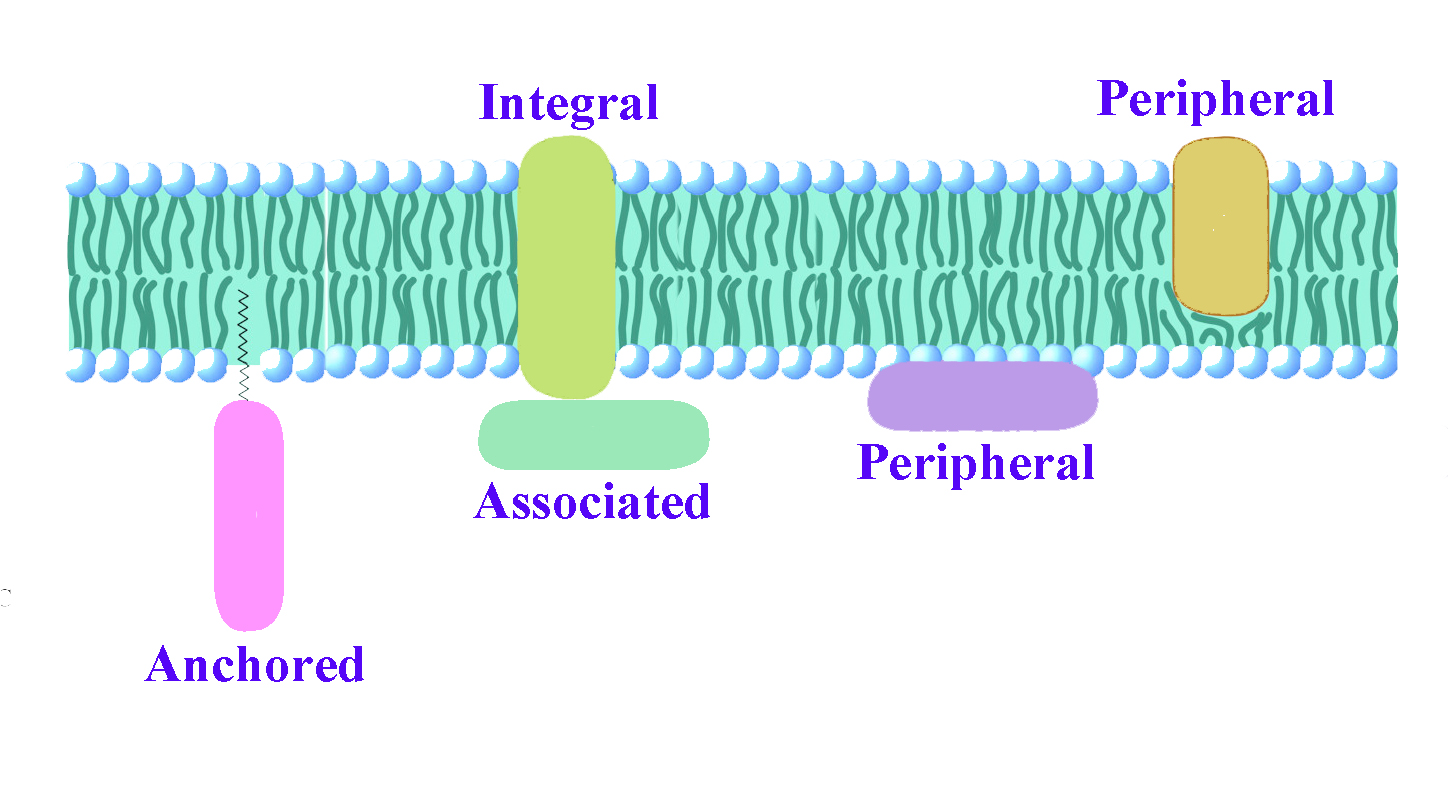
Figura\(\PageIndex{10}\): Tipos de proteínas de membrana
La geometría de la bicapa lipídica es tal que es hidrofóbica en su interior e hidrófila en el exterior. Tales propiedades también dictan las cadenas laterales de aminoácidos de las proteínas que interactúan con la bicapa. Para la mayoría de las proteínas de membrana, los aminoácidos polares se encuentran donde la proteína se proyecta a través de la bicapa (interactuando con sustancias acuosas/polares) y los aminoácidos no polares se incrustan dentro de la porción no polar de la bicapa que contiene las colas de ácidos grasos.Los glicolípidos y las glicoproteínas juegan importante roles en la identificación celular. Los tipos sanguíneos, por ejemplo, difieren entre sí en la estructura de las cadenas de carbohidratos que sobresalen de la superficie de la glicoproteína en sus membranas.Las células tienen cientos de proteínas de membrana y la composición proteica de una membrana varía con su función y ubicación. Las membranas mitocondriales se encuentran entre las más densamente empaquetadas con proteínas. La membrana plasmática tiene un gran número de proteínas integrales involucradas en la comunicación de información a través de la membrana (señalización) o en el transporte de materiales hacia la célula.
Transporte de Membrana
Los materiales, como los alimentos y los desechos, deben moverse a través de la bicapa lipídica de una célula. Hay dos medios para lograrlo: los procesos pasivos y los procesos activos. Los procesos pasivos tienen como única fuerza impulsora el proceso de difusión. En estos sistemas, las moléculas siempre pasan de una concentración mayor a una concentración menor. Estos pueden ocurrir directamente a través de una membrana (agua, oxígeno, dióxido de carbono y monóxido de carbono) o a través de proteínas de transporte especiales (proteínas de transporte de glucosa de los glóbulos rojos, por ejemplo). En cada caso, no se gasta energía celular en el movimiento de las moléculas. Por otro lado, los procesos activos requieren energía para lograr dicho transporte. Una fuente de energía común es ATP (ver Na+/K+ ATPasa), pero se emplean muchas otras fuentes de energía. Por ejemplo, el transportador de sodio-glucosa utiliza un gradiente de sodio como fuerza para transportar activamente la glucosa a una célula. Por lo tanto, es importante saber que no todo el transporte activo utiliza energía ATP. Las proteínas, como el transportador de sodio-glucosa que mueven dos moléculas en la misma dirección a través de la membrana se llaman simporters (también llamados synporters). Si la acción de una proteína en el movimiento de iones a través de una membrana da como resultado un cambio en la carga, la proteína se describe como electrogénica y si no hay cambio en la carga la proteína se describe como electro-neutra.
Na+/K+ ATPasa
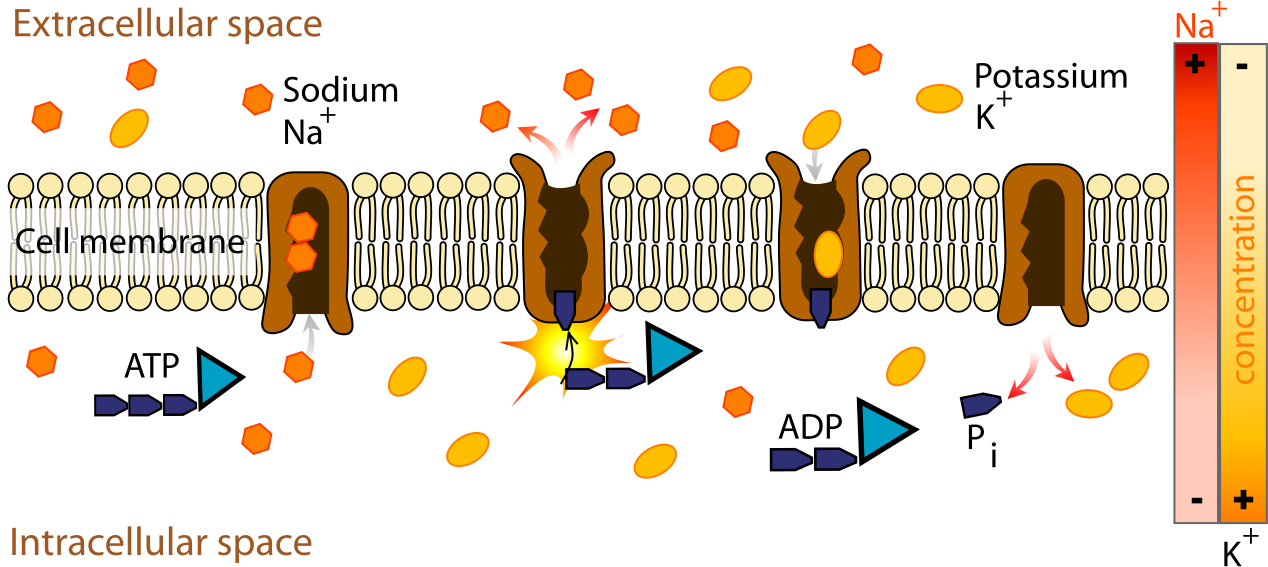
Figura\(\PageIndex{11}\): ATPasa de sodio y potasio
Otra proteína integral importante de la membrana es la Na+/K+ ATPasa, que transporta iones de sodio fuera de la célula e iones de potasio a la célula. La proteína, que se describe como un anti-puerto (moléculas movidas en direcciones opuestas a través de la membrana) utiliza la energía del ATP para crear gradientes iónicos que son importantes tanto para mantener la presión osmótica celular como (en las células nerviosas) para crear los gradientes iónicos necesarios para la transmisión de la señal. El sistema de transporte mueve tres átomos de sodio fuera de la célula y dos átomos de potasio hacia la célula por cada ATP hidrolizado.
Bacteriorodopsina
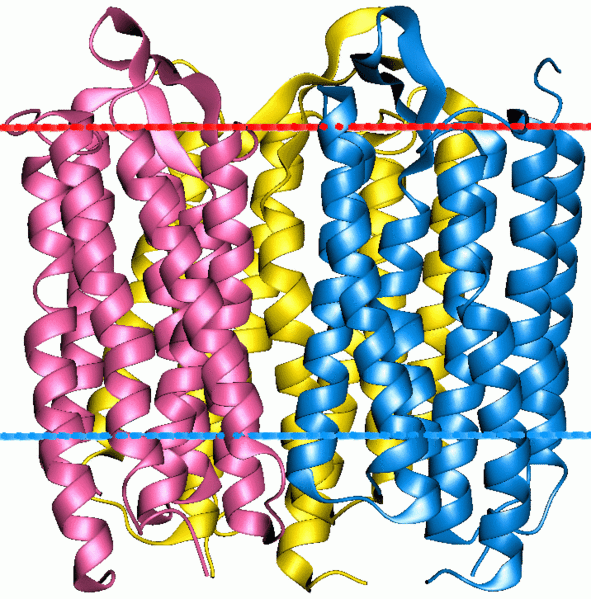
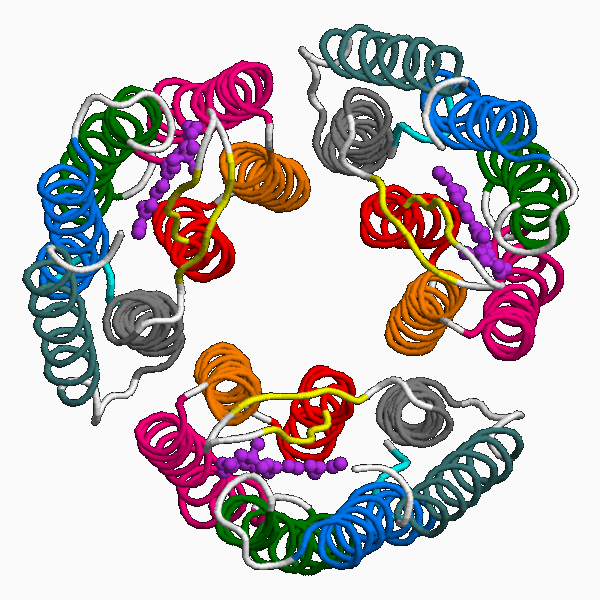
Una proteína de membrana integral interesante es la bacteriorodopsina. La proteína tiene tres cadenas polipeptídicas idénticas, cada una girada 120 grados con respecto a las otras. Cada cadena tiene siete hélices alfa transmembrana y contiene una molécula de retina (Vitamina A) enterrada profundamente dentro de cada cavidad (mostrada en púrpura en la figura inferior a la izquierda). La vitamina A es sensible a la luz y se isomeriza rápidamente entre una forma cis y una trans en presencia de luz. La conformación cambiante de la vitamina A se utiliza para transportar protones a través de la proteína y fuera de la bacteria, creando un gradiente de protones a través de la membrana celular, que se usa en última instancia para producir ATP. No es demasiado difícil imaginar la ingeniería de un organismo (digamos un pez transparente) para que contenga bacteriorodopsina en su membrana interna mitocondrial. Cuando la luz brilla sobre ella, la bacteriorodopsina podría usarse para generar un gradiente de protones (al igual que lo hace el transporte de electrones) y potenciar la fosforilación oxidativa. Tal pez sería en parte fotosintético ya que estaría derivando energía de la luz, pero diferiría de las plantas en ser incapaz de asimilar el dióxido de carbono en una serie de “reacciones oscuras”.
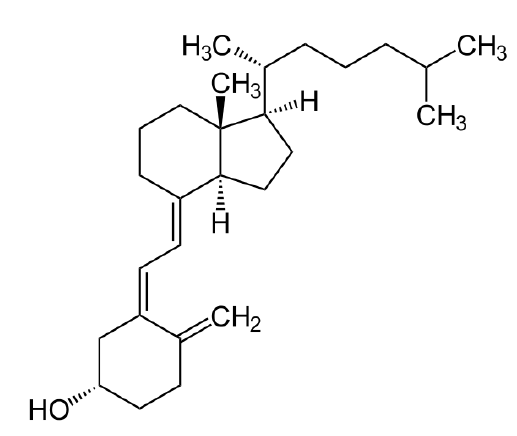
Figura\(\PageIndex{13}\): Vitamin D
Vitaminas liposolubles
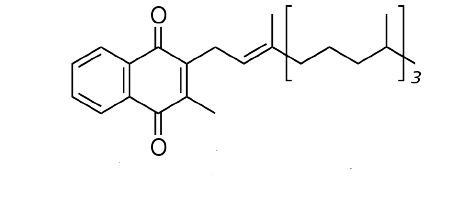 Figura\(\PageIndex{1}\): Vitamina K
Figura\(\PageIndex{1}\): Vitamina K
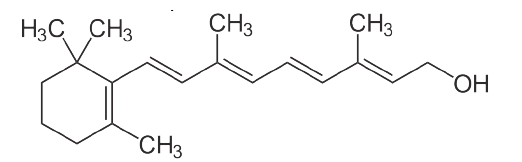
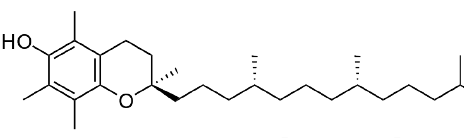
Otros lípidos destacados incluyen las vitaminas liposolubles - A, D, E y K. La vitamina A viene en tres formas químicas primarias, retinol (almacenamiento en el hígado), retinal (papel en la visión) y ácido retinoico (papeles en el crecimiento y desarrollo). La vitamina D (colecalciferol) juega un papel importante en la absorción intestinal de calcio y fosfato y por lo tanto en huesos sanos. Derivado en última instancia del colesterol, el compuesto se puede sintetizar en una reacción catalizada por la luz ultravioleta. La vitamina E (tocoferol) es la vitamina de la que menos se conoce. Consta de un grupo de ocho compuestos liposolubles de los cuales el isómero alfa tiene la mayor actividad biológica. La vitamina K (el nombre proviene del alemán para la vitamina de coagulación) es esencial para la coagulación de la sangre. Se utiliza como cofactor para la enzima que modifica la protrombina para aumentar su afinidad por el calcio, permitiendo que se posicione más cerca del sitio de una herida.