
6.1.1.4: Cadenas Alimentarias y Redes Alimentarias
- Page ID
- 54378

Las interacciones tróficas en una comunidad pueden ser representadas por diagramas llamados cadenas alimentarias y redes alimentarias. Antes de discutir estas representaciones en detalle, primero debemos revisar los fundamentos de la energía. La energía fluye a través de una comunidad como resultado de interacciones tróficas.
Energía
Prácticamente todas las tareas que realizan los organismos vivos requieren energía. En general, la energía se define como la capacidad de hacer trabajo, o de crear algún tipo de cambio. La energía existe en diferentes formas. Los ejemplos incluyen la energía luminosa, la energía cinética, la energía térmica, la energía potencial y la energía química.
Cuando un objeto está en movimiento, hay energía asociada a ese objeto. Piensa en una bola de naufragio. Incluso una bola de naufragio de movimiento lento puede hacer mucho daño a otros objetos. La energía asociada con los objetos en movimiento se llama energía cinética. La energía térmica es la energía del movimiento en la materia (cualquier cosa que ocupe espacio y tenga masa) y se considera un tipo de energía cinética. Cuanto más caliente es la sustancia, más rápido se mueven sus moléculas. El rápido movimiento de las moléculas en el aire, una bala a toda velocidad y una persona que camina tienen todos energía cinética. Ahora, ¿y si esa misma bola de naufragio inmóvil se levanta dos pisos sobre el suelo con una grúa? Si la bola de naufragio suspendida no se mueve, ¿hay energía asociada a ella? La respuesta es sí. La energía que se requería para levantar la bola de naufragio no desapareció, sino que ahora se almacena en la bola de naufragio en virtud de su posición y la fuerza de gravedad que actúa sobre ella. Este tipo de energía se llama energía potencial. Si la pelota cayera, la energía potencial se transformaría en energía cinética hasta que se agotara toda la energía potencial cuando la pelota descansara en el suelo. Las bolas de naufragio también se balancean como un péndulo; a través del columpio, hay un cambio constante de energía potencial (más alta en la parte superior del columpio) a energía cinética (más alta en la parte inferior del columpio). Otros ejemplos de energía potencial incluyen la energía del agua contenida detrás de una presa o una persona a punto de lanzarse en paracaidismo fuera de un avión (figura\(\PageIndex{a}\)).
La energía potencial no sólo está asociada con la ubicación de la materia, sino también con la estructura de la materia. La energía química es un ejemplo de energía potencial que se almacena en moléculas. Cuando las moléculas que son de mayor energía y menos estables reaccionan para formar productos que son de menor energía y más estables, esta energía almacenada se libera. La energía química es responsable de proporcionar a las células vivas energía de los alimentos.
Para apreciar la forma en que la energía fluye dentro y fuera de los sistemas biológicos, es importante comprender dos de las leyes físicas que rigen la energía. La primera ley de la termodinámica establece que la cantidad total de energía en el universo es constante y conservada. Es decir, siempre ha habido, y siempre habrá, exactamente la misma cantidad de energía en el universo. La energía existe en muchas formas diferentes. Según la primera ley de la termodinámica, la energía puede transferirse de un lugar a otro o transformarse en diferentes formas, pero no puede crearse ni destruirse. Las transferencias y transformaciones de la energía ocurren a nuestro alrededor todo el tiempo. Las bombillas transforman la energía eléctrica en energía lumínica y térmica. Las estufas de gas transforman la energía química del gas natural en energía térmica. Las plantas realizan una de las transformaciones energéticas más útiles biológicamente en la tierra: la de convertir la energía de la luz solar en energía química almacenada dentro de moléculas biológicas, como los azúcares (figura\(\PageIndex{b}\)).

El reto para todos los organismos vivos es obtener energía de su entorno en formas que sean utilizables para realizar trabajos celulares. Las tareas primarias de una célula viva de obtener, transformar y usar energía para hacer el trabajo pueden parecer simples. No obstante, la segunda ley de la termodinámica explica por qué estas tareas son más difíciles de lo que parecen. Todas las transferencias y transformaciones de energía nunca son completamente eficientes. En cada transferencia de energía, se pierde cierta cantidad de energía en una forma que es inutilizable. En la mayoría de los casos, esta forma es energía térmica. Por ejemplo, cuando se enciende una bombilla, parte de la energía que se convierte de energía eléctrica en energía lumínica se pierde como energía térmica. De igual manera, se pierde algo de energía como energía térmica durante las reacciones metabólicas que ocurren en los organismos.
El concepto de orden y desorden se relaciona con la segunda ley de la termodinámica. Cuanta más energía pierde un sistema a su entorno, menos ordenado y más aleatorio es el sistema. Los científicos se refieren a la medida de aleatoriedad o trastorno dentro de un sistema como entropía. Alta entropía significa trastorno alto y baja energía. Los seres vivos están altamente ordenados, requiriendo un aporte de energía constante para mantenerse en un estado de baja entropía.
Flujo de Energía
Las células funcionan con la energía química que se encuentra principalmente en las moléculas de carbohidratos, y la mayoría de estas moléculas son producidas por un solo proceso: la fotosíntesis. A través de la fotosíntesis, ciertos organismos convierten la energía solar (luz solar) en energía química, que luego se utiliza para construir moléculas de carbohidratos (figura\(\PageIndex{c}\)). La energía que se aprovecha de la fotosíntesis ingresa continuamente a las comunidades y se transfiere de un organismo a otro. Por lo tanto, directa o indirectamente, el proceso de fotosíntesis proporciona la mayor parte de la energía que requieren los seres vivos en la Tierra. Consulta el Ciclo del Carbono y la Fotosíntesis en OpenStax Concepts of Biology para más detalles sobre la fotosíntesis.

Los organismos que realizan la fotosíntesis (como plantas, algas y algunas bacterias), y los organismos que sintetizan azúcares a través de otros medios se denominan productores. Sin estos organismos, la energía no estaría disponible para otros organismos vivos, y la vida no sería posible. Los consumidores, como animales, hongos y diversos microorganismos dependen de los productores, ya sea directa o indirectamente. Por ejemplo, un venado obtiene energía al comer plantas. Un lobo que se come un venado obtiene energía que originalmente provenía de las plantas que comía ese venado (figura\(\PageIndex{d}\)). A partir de este razonamiento, todos los alimentos consumidos por los humanos se remontan a productores que realizan la fotosíntesis (figura\(\PageIndex{e}\)).
Los consumidores pueden clasificarse en función de si comen material animal o vegetal (figura\(\PageIndex{f}\)). Los consumidores que se alimentan exclusivamente de animales se llaman carnívoros. Leones, tigres, serpientes, tiburones, estrellas de mar, arañas y mariquitas son todos carnívoros. Los herbívoros son consumidores que se alimentan exclusivamente de material vegetal, y ejemplos incluyen ciervos, koalas, algunas especies de aves, grillos y orugas. Los herbívoros pueden clasificarse además en frugívoros (comedores de frutas), granívoros (comedores de semillas), nectivores (alimentadores de néctar) y folivoros (comedores de hojas). Los consumidores que comen tanto material vegetal como animal son considerados omnívoros. Humanos, osos, pollos, cucarachas y cangrejos de río son ejemplos de omnívoros.
Los productores y consumidores muertos son consumidos por detritivores (que ingieren tejidos muertos) y descomponedores (que descomponen aún más estos tejidos en moléculas simples al secretar enzimas digestivas). Los animales invertebrados, como gusanos y milpiés, son ejemplos de detritívoros, mientras que los hongos y ciertas bacterias son ejemplos de descomponedores.
Cadenas Alimentarias
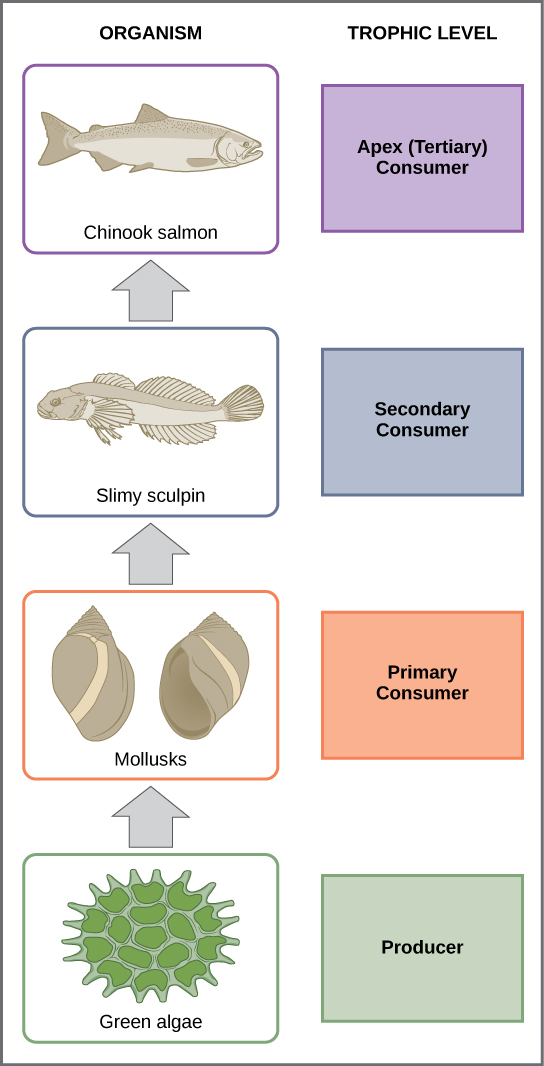
Una cadena alimentaria es una secuencia lineal de organismos a través de los cuales los nutrientes y la energía pasan a medida que un organismo se come a otro. Cada organismo en una cadena alimentaria ocupa un nivel trófico específico (nivel de energía), su posición en la cadena alimentaria. El primer nivel trófico en la cadena alimentaria son los productores. Los consumidores primarios (los herbívoros que comen productores) son el segundo nivel trófico. A continuación están los consumidores de nivel superior. Los consumidores de nivel superior incluyen consumidores secundarios (tercer nivel trófico), que suelen ser carnívoros que comen a los consumidores primarios, y consumidores terciarios (cuarto nivel trófico), que son carnívoros que comen a otros carnívoros. En la cadena alimentaria del lago Ontario, que se muestra en la figura\(\PageIndex{g}\), el salmón Chinook es el consumidor ápice en la parte superior de esta cadena alimentaria. Algunas comunidades tienen niveles tróficos adicionales (consumidores cuaternarios, consumidores de quinto orden, etc.). Finalmente, los detrívoros y descomponedores descomponen los organismos muertos y en descomposición desde cualquier nivel trófico. Hay un solo camino a través de una cadena alimentaria.
Un factor importante que limita el número de pasos en una cadena alimentaria es la energía. Solo alrededor del 10% de la energía en un nivel trófico se transfiere al siguiente nivel trófico. Esto se debe a que mucha energía se pierde como calor durante las transferencias entre niveles tróficos o a descomponedores debido a la segunda ley de la termodinámica. Así, después de cuatro a seis transferencias tróficas de energía, la cantidad de energía que queda en la cadena alimentaria puede no ser lo suficientemente grande como para apoyar a poblaciones viables en niveles tróficos más altos (ver también Productividad Comunitaria y Eficiencia de Transferencia).
Ciertas toxinas ambientales pueden concentrarse más a medida que avanzan en la cadena alimentaria, con las concentraciones más altas que ocurren en los principales consumidores, un proceso llamado biomagnificación. Esencialmente, un consumidor superior ingiere todas las toxinas que previamente se habían acumulado en los cuerpos de los organismos en los niveles tróficos más bajos. Esto explica por qué comer frecuentemente ciertos peces, como el atún o el pez espada, aumenta tu exposición al mercurio, un metal pesado tóxico.
Webs Alimentarias
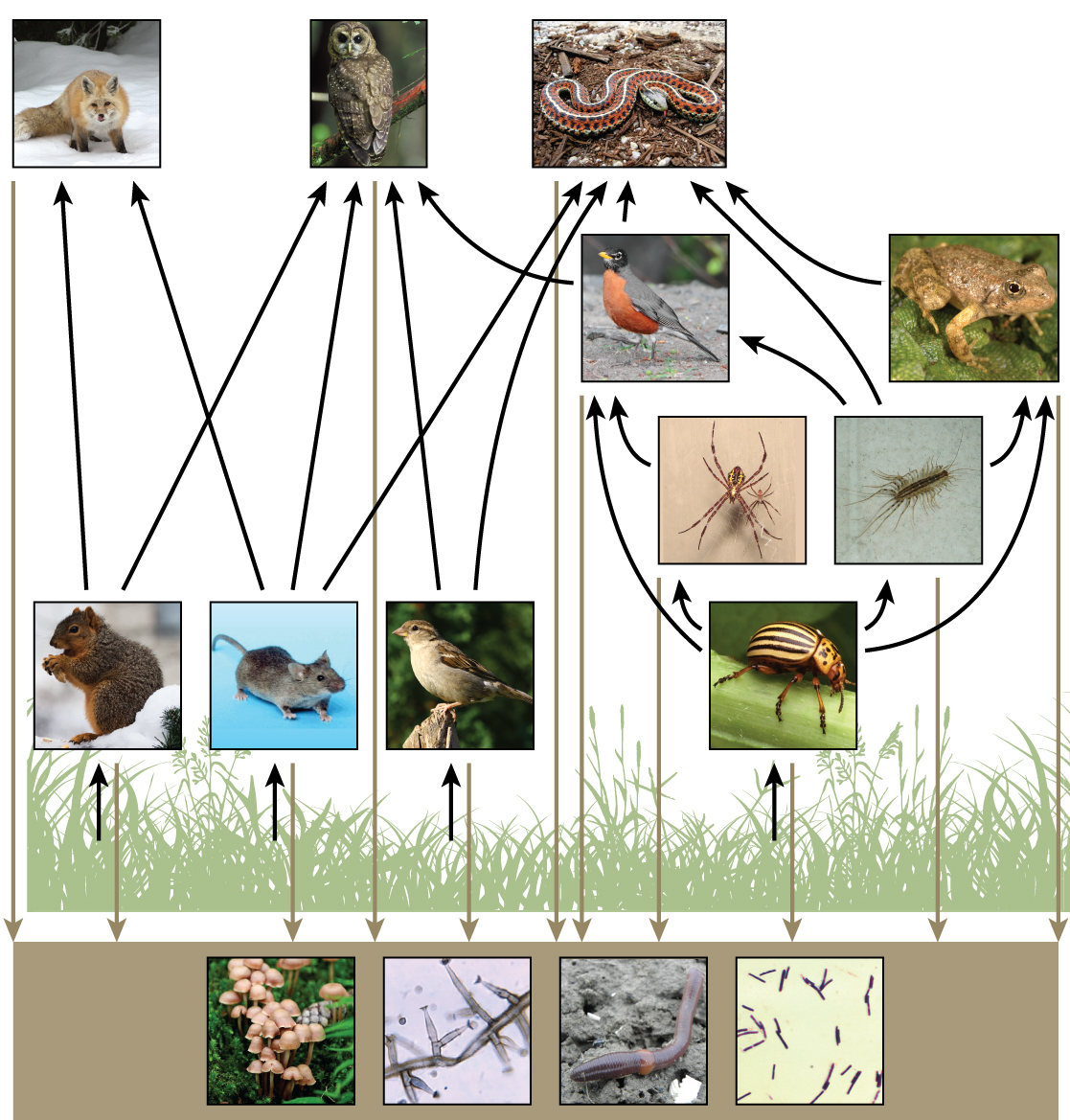
Si bien las cadenas alimentarias son simples y fáciles de analizar, existe un problema al usar las cadenas alimentarias para describir a la mayoría de las comunidades. Incluso cuando todos los organismos se agrupan en niveles tróficos apropiados, algunos de estos organismos pueden alimentarse a más de un nivel trófico. Además, las especies se alimentan y son consumidas por más de una especie. En otras palabras, el modelo lineal de interacciones tróficas, la cadena alimentaria, es una representación hipotética y excesivamente simplista de la estructura comunitaria. Un modelo holístico, que incluye todas las interacciones entre diferentes especies y sus complejas relaciones interconectadas entre sí y con el medio ambiente, es un modelo más preciso y descriptivo. Una red alimentaria es un concepto que da cuenta de las múltiples interacciones tróficas entre cada especie (figura\(\PageIndex{h}\) e i).
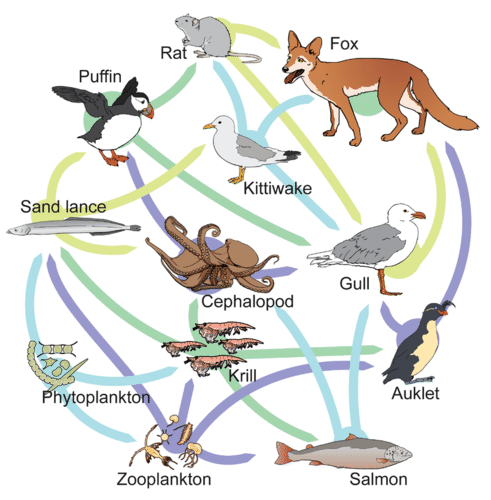
El nivel trófico de cada especie en una red alimentaria no es necesariamente un número entero. En la figura\(\PageIndex{i}\), el fitoplancton son los productores primarios (nivel trófico 1). El zooplancton solo se alimenta de fitoplancton, convirtiéndolos en consumidores primarios (nivel trófico 2). Determinar el nivel trófico de las otras especies es más complejo. Por ejemplo, el krill come tanto fitoplancton como zooplancton. Si el krill solo comiera fitoplancton serían consumidores primarios (nivel trófico 2). Si comieran solo zooplancton, serían consumidores secundarios (nivel trófico 3). Ya que, el krill consume ambos, su nivel trófico es 2.5.
Productividad comunitaria y eficiencia de transferencia
La tasa a la que los productores fotosintéticos incorporan energía del sol se denomina productividad primaria bruta. En un pantano de totora, las plantas solo atrapan 2.2% de la energía del sol que las alcanza. El tres por ciento de la energía se refleja, y otro 94.8% se utiliza para calentar y evaporar el agua dentro y alrededor de la planta. Sin embargo, no toda la energía incorporada por los productores está disponible para los demás organismos en la red alimentaria porque los productores también deben crecer y reproducirse, lo que consume energía. Al menos la mitad del 2.2% atrapado por las plantas de pantano de espadaña se utiliza para satisfacer las necesidades energéticas propias de las plantas.
La productividad primaria neta es la energía que permanece en los productores después de contabilizar las necesidades metabólicas de los productores y la pérdida de calor. La productividad neta está entonces disponible para los consumidores primarios en el siguiente nivel trófico. Una manera de medir la productividad primaria neta es recolectar y pesar el material vegetal producido en un m 2 (aproximadamente 10.7 pies 2) de tierra en un intervalo dado. Un gramo de material vegetal (por ejemplo, tallos y hojas), que es en gran parte la celulosa carbohidrato, produce alrededor de 4.25 kcal de energía cuando se quema. La productividad primaria neta puede variar de 500 kcal/m 2 /año en el desierto a 15,000 kcal/m 2 /año en una selva tropical.
En una comunidad acuática de Silver Springs, Florida, la productividad primaria bruta (energía total acumulada por los productores primarios) fue de 20,810 kcal/m 2 /año (cifra\(\PageIndex{j}\)). La productividad primaria neta (energía disponible para los consumidores) fue de solo 7,632 kcal/m 2 /año después de contabilizar la pérdida de energía como el calor y la energía requieren para satisfacer las necesidades metabólicas del productor.
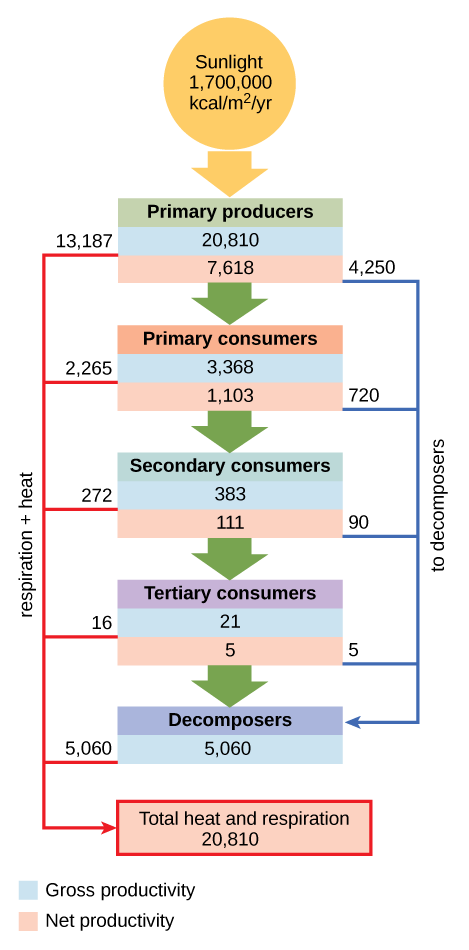
Solo una fracción de la energía capturada por un nivel trófico es asimilada a biomasa, lo que la pone a disposición del siguiente nivel trófico. La asimilación es la biomasa del nivel trófico actual después de contabilizar la energía perdida por ingestión incompleta de alimentos, la energía utilizada para realizar el trabajo por ese nivel trófico y la energía perdida como desperdicio. La ingestión incompleta se refiere al hecho de que algunos consumidores comen solo una parte de sus alimentos. Por ejemplo, cuando un león mata a un antílope, se lo comerá de todo excepto la piel y los huesos. Al león le falta la médula ósea rica en energía dentro del hueso, por lo que el león no hace uso de todas las calorías que su presa podría proporcionar. En Silver Springs, solo 1103 kcal/m 2 /año de los 7618 kcal/m 2 /año de energía disponible para los consumidores primarios fueron asimilados a su biomasa. (La eficiencia de transferencia de nivel trófico entre los dos primeros niveles tróficos fue de aproximadamente 14.8 por ciento).
La fuente de calor de un animal influye en sus necesidades energéticas. Los ectotermos, como invertebrados, peces, anfibios y reptiles, dependen de fuentes externas para el calor corporal, y las endotermas, como aves y mamíferos, dependen del calor generado internamente. Generalmente, los ectotermos requieren menos energía para satisfacer sus necesidades metabólicas y que las endotermas, y por lo tanto, muchas endotermas tienen que comer con más frecuencia que los ectotermos.
La ineficiencia del uso de energía por parte de las endotermas tiene amplias implicaciones para el suministro mundial de alimentos. Es ampliamente aceptado que la industria cárnica utiliza grandes cantidades de cultivos para alimentar al ganado, y debido a que un bajo porcentaje de esta se asimila a la biomasa, gran parte de la energía de la alimentación animal se pierde. Por ejemplo, cuesta alrededor de 1¢ producir 1000 calorías dietéticas (kcal) de maíz o soja, pero aproximadamente $0.19 producir un número similar de calorías cultivando ganado para el consumo de carne. El mismo contenido energético de la leche del ganado bovino también es costoso, a aproximadamente $0.16 por 1000 kcal. Así, ha habido un movimiento creciente a nivel mundial para promover el consumo de alimentos no cárnicos y no lácteos para que se desperdicie menos energía alimentando animales para la industria cárnica.
Atribución
Modificado por Melissa Ha de las siguientes fuentes:
- La energía, la energía ingresa a los ecosistemas a través de la fotosíntesis y el flujo de energía a través de los ecosistemas desde la biología ambiental por Matthew R. Fisher (con licencia CC-BY
- Ecología de Ecosistemas, Flujo de Energía a través de Ecosistemas y Sistemas Digestivos de Biología General/Biología 2e por OpenStax (licenciado bajo CC-BY). Accede gratis en openstax.org.
- Productividad de Ecosistemas a partir de la Biología por John W. Kimball (licenciado bajo CC-BY)