
8.6: La fotosíntesis y la importancia de la luz
- Page ID
- 54694

Objetivos de aprendizaje
- Describir la función y ubicación de los pigmentos fotosintéticos en eucariotas y procariotas
- Describir los principales productos de las reacciones dependientes de la luz e independientes de la luz
- Describir las reacciones que producen glucosa en una célula fotosintética
- Comparar y contrastar fotofosforilación cíclica y no cíclica
Los organismos heterótrofos que van desde E. coli hasta humanos dependen de la energía química que se encuentra principalmente en las moléculas de carbohidratos. Muchos de estos carbohidratos son producidos por la fotosíntesis, el proceso bioquímico por el cual los organismos fototróficos convierten la energía solar (luz solar) en energía química. Aunque la fotosíntesis se asocia más comúnmente con las plantas, la fotosíntesis microbiana también es un importante proveedor de energía química, alimentando muchos ecosistemas diversos. En esta sección, nos centraremos en la fotosíntesis microbiana.
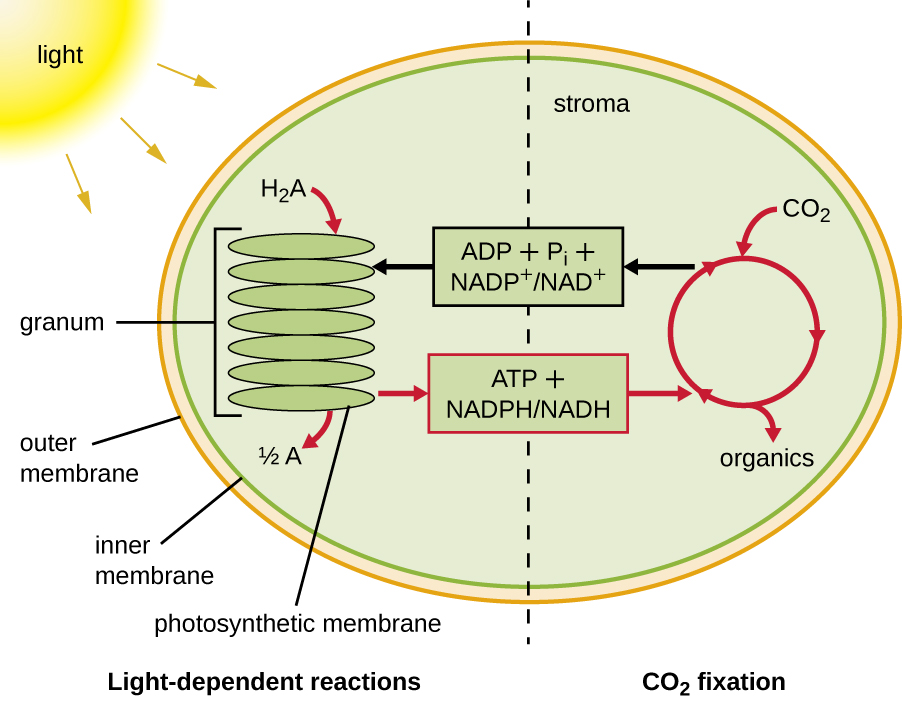
La fotosíntesis se realiza en dos etapas secuenciales: las reacciones dependientes de la luz y las reacciones independientes de la luz (Figura\(\PageIndex{1}\)). En las reacciones dependientes de la luz, la energía de la luz solar es absorbida por moléculas de pigmento en membranas fotosintéticas y convertida en energía química almacenada. En las reacciones independientes de la luz, el ener gy químico producido por las reacciones dependientes de la luz se utiliza para impulsar el ensamblaje de moléculas de azúcar usando CO 2; sin embargo, estas reacciones siguen dependiendo de la luz porque los productos de las reacciones dependientes de la luz necesarios para conducirlas son cortos -vivido. Las reacciones dependientes de la luz producen ATP y NADPH o NADH para almacenar energía temporalmente. Estos portadores de energía se utilizan en las reacciones independientes de la luz para impulsar el proceso energéticamente desfavorable de “fijar” CO 2 inorgánico en una forma orgánica, el azúcar.
Estructuras fotosintéticas en eucariotas y procariotas
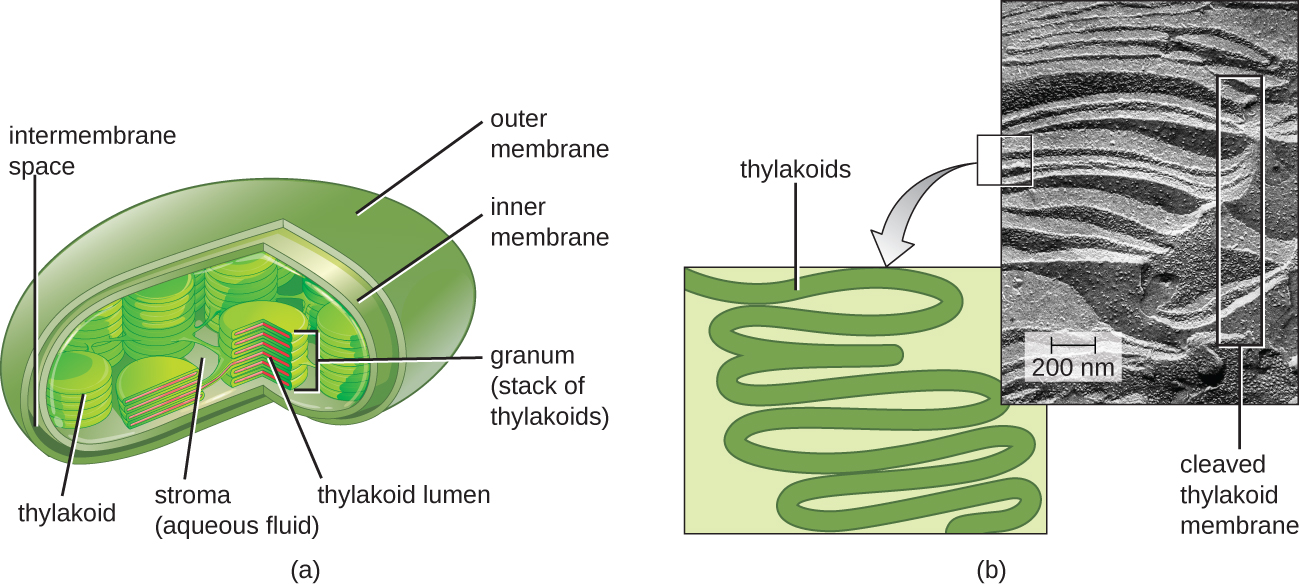
En todos los eucariotas fototróficos, la fotosíntesis se realiza dentro de un cloroplasto, un orgánulo que surgió en eucariotas por endosimbiosis de una bacteria fotosintética (ver Características únicas de las células eucariotas). Estos cloroplastos están encerrados por una doble membrana con capas interna y externa. Dentro del cloroplasto se encuentra una tercera membrana que forma estructuras fotosintéticas apiladas en forma de disco llamadas tilacoides (Figura\(\PageIndex{2}\)). Una pila de tilacoides se llama gránulo, y el espacio que rodea al gránulo dentro del cloroplasto se llama estroma.
Las membranas fotosintéticas en procariotas, por el contrario, no están organizadas en distintos orgánulos encerrados en membranas; más bien, son regiones plegadas de la membrana plasmática. En las cianobacterias, por ejemplo, estas regiones plegadas también se denominan tilacoides. En cualquier caso, incrustadas dentro de las membranas tilacoides u otras membranas bacterianas fotosintéticas se encuentran moléculas pigmentarias fotosintéticas organizadas en uno o más fotosistemas, donde la energía luminosa se convierte realmente en energía química.
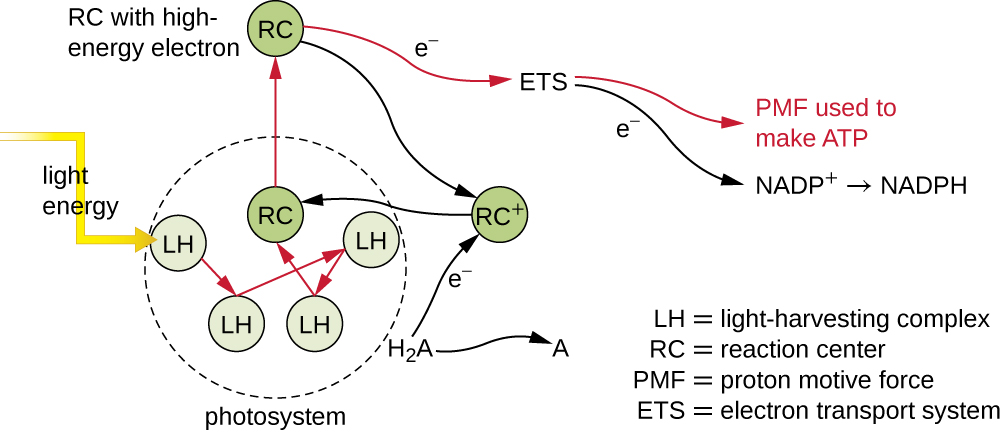
Los pigmentos fotosintéticos dentro de las membranas fotosintéticas se organizan en fotosistemas, cada uno de los cuales está compuesto por un complejo de recolección de luz (antenas) y un centro de reacción. El complejo de recolección de luz consiste en múltiples proteínas y pigmentos asociados que cada uno puede absorber la energía de la luz y, así, excitarse. Esta energía se transfiere de una molécula de pigmento a otra hasta que finalmente (después de aproximadamente una millonésima parte de segundo) se entrega al centro de reacción. Hasta este punto, solo la energía, no los electrones, se ha transferido entre moléculas. El centro de reacción contiene una molécula de pigmento que puede sufrir oxidación tras la excitación, en realidad renunciando a un electrón. Es en este paso de la fotosíntesis que la energía luminosa se convierte en un electrón excitado.
Diferentes tipos de pigmentos de recolección de luz absorben patrones únicos de longitudes de onda (colores) de luz visible. Los pigmentos reflejan o transmiten las longitudes de onda que no pueden absorber, haciéndolos aparecer del color correspondiente. Ejemplos de pigmentos fotosintéticos (moléculas utilizadas para absorber energía solar) son bacterioclorofilas (verdes, moradas o rojas), carotenoides (naranja, rojo o amarillo), clorofilas (verde), ficocianinas (azul) y ficoeritrinas (rojo). Al tener mezclas de pigmentos, un organismo puede absorber energía de más longitudes de onda. Debido a que las bacterias fotosintéticas comúnmente crecen en competencia por la luz solar, cada tipo de bacteria fotosintética está optimizada para cosechar las longitudes de onda de la luz a las que comúnmente se expone, lo que lleva a la estratificación de comunidades microbianas en ecosistemas acuáticos y de suelo por calidad de luz y penetración.
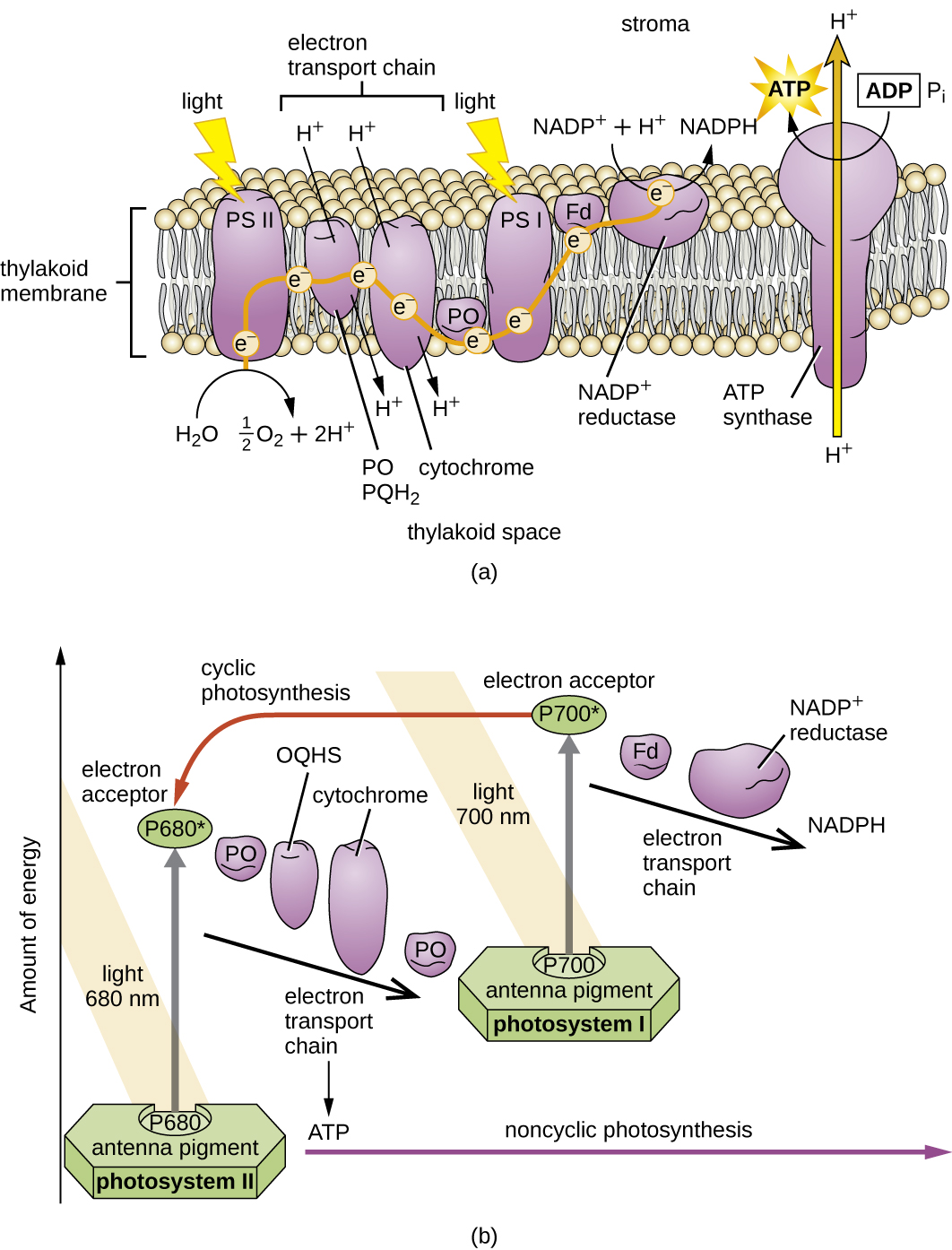
Una vez que el complejo de captación de luz transfiere la energía al centro de reacción, el centro de reacción entrega sus electrones de alta energía, uno por uno, a un portador de electrones en un sistema de transporte de electrones, y se inicia la transferencia de electrones a través del ETS. El ETS es similar al utilizado en la respiración celular y está incrustado dentro de la membrana fotosintética. En última instancia, el electrón se utiliza para producir NADH o NADPH. El gradiente electroquímico que se forma a través de la membrana fotosintética se utiliza para generar ATP por quimioósmosis a través del proceso de fotofosforilación, otro ejemplo de fosforilación oxidativa (Figura\(\PageIndex{3}\)).
Ejercicio\(\PageIndex{1}\)
En un eucariota fototrófico, ¿dónde tiene lugar la fotosíntesis?
Fotosíntesis Oxígena y Anoxigénica
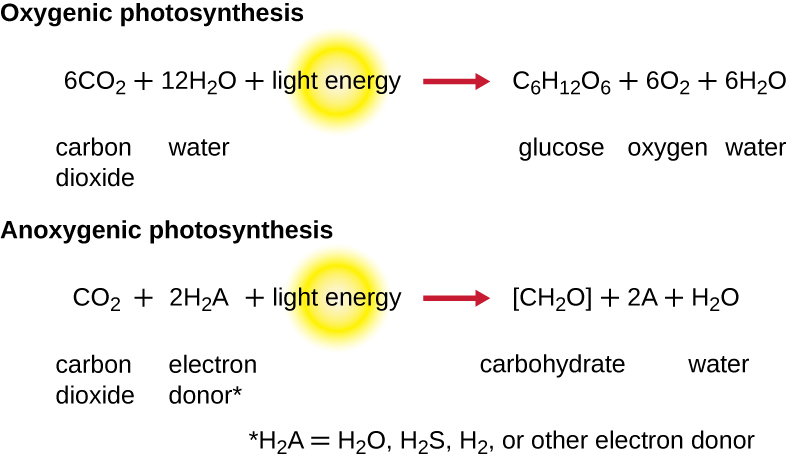
Para que la fotosíntesis continúe, se debe reemplazar el electrón perdido del pigmento del centro de reacción. La fuente de este electrón (H 2 A) diferencia la fotosíntesis oxigénica de plantas y cianobacterias de la fotosíntesis anoxígena realizada por otros tipos de fototrofos bacterianos (Figura\(\PageIndex{4}\)). En la fotosíntesis oxigénica, H 2 O se divide y suministra el electrón al centro de reacción. Debido a que el oxígeno se genera como subproducto y se libera, este tipo de fotosíntesis se conoce como fotosíntesis oxigénica. Sin embargo, cuando otros compuestos reducidos sirven como donadores de electrones, no se genera oxígeno; estos tipos de fotosíntesis se denominan fotosíntesis anoxígena. El sulfuro de hidrógeno (H 2 S) o el tiosulfato\(\ce{(S2O3^{2-})}\) pueden servir como donador de electrones, generando\(\ce{(SO4^{2-})}\) iones de azufre elemental y sulfato, respectivamente, como resultado.
Los fotosistemas se han clasificado en dos tipos: fotosistema I (PSI) y fotosistema II (PSII) (Figura\(\PageIndex{5}\)). Las cianobacterias y los cloroplastos vegetales tienen ambos fotosistemas, mientras que las bacterias fotosintéticas anoxigénicas utilizan solo uno de los fotosistemas. Ambos fotosistemas son excitados por la energía de la luz simultáneamente. Si la célula requiere tanto ATP como NADPH para la biosíntesis, entonces realizará fotofosforilación no cíclica. Al pasar el electrón del centro de reacción PSII al ETS que conecta PSII y PSI, el electrón perdido del centro de reacción PSII es reemplazado por la división del agua. El electrón del centro de reacción PSI excitado se utiliza para reducir el NADP + a NADPH y es reemplazado por el electrón que sale del ETS. El flujo de electrones de esta manera se llama el esquema Z.
Si la necesidad de ATP de una célula es significativamente mayor que su necesidad de NADPH, puede evitar la producción de energía reductora a través de la fotofosforilación cíclica. Solo se usa PSI durante la fotofosforilación cíclica; el electrón de alta energía del centro de reacción PSI se pasa a un portador ETS y luego finalmente regresa al pigmento del centro de reacción PSI oxidado, reduciéndolo así.
Ejercicio\(\PageIndex{2}\)
¿Por qué una bacteria fotosintética tendría diferentes pigmentos?
Reacciones independientes de la luz
Después de que la energía del sol se convierte en energía química y se almacena temporalmente en moléculas de ATP y NADPH (que tienen una vida útil de millonésimas de segundo), los fotoautótrofos tienen el combustible necesario para construir moléculas de carbohidratos multicarbon, que pueden sobrevivir durante cientos de millones de años, a largo plazo almacenamiento de energía. El carbono proviene del CO 2, el gas que es un producto de desecho de la respiración celular.
El ciclo Calvin-Benson (llamado así por Melvin Calvin [1911—1997] y Andrew Benson [1917—2015]), la vía bioquímica utilizada para la fijación del CO 2, se localiza dentro del citoplasma de bacterias fotosintéticas y en el estroma de cloroplastos eucariotas. Las reacciones independientes de la luz del ciclo Calvino se pueden organizar en tres etapas básicas: fijación, reducción y regeneración (ver Apéndice C para una ilustración detallada del ciclo Calvino).
- Fijación: La enzima ribulosa bisfosfato carboxilasa (RuBisCO) cataliza la adición de CO 2 al bifosfato de ribulosa (RuBP). Esto da como resultado la producción de 3-fosfoglicerato (3-PGA).
- Reducción: Se utilizan seis moléculas de ATP y NADPH (de las reacciones dependientes de la luz) para convertir 3-PGA en gliceraldehído 3-fosfato (G3P). Luego se usa algo de G3P para generar glucosa.
- Regeneración: El G3P restante no utilizado para sintetizar glucosa se usa para regenerar RuBP, permitiendo que el sistema continúe con la fijación de CO 2. En estas reacciones de regeneración se utilizan tres moléculas más de ATP.
El ciclo Calvin es ampliamente utilizado por plantas y bacterias fotoautótrofas, y se dice que la enzima RubiSco es la enzima más abundante en la tierra, al componer 30% — 50% de la proteína soluble total en cloroplastos vegetales. 1 Sin embargo, además de su uso prevalente en fotoautótrofos, el ciclo Calvino también es utilizado por muchos quimioautótrofos no fotosintéticos para fijar CO 2. Adicionalmente, otras bacterias y arqueas utilizan sistemas alternativos para la fijación de CO 2. Aunque la mayoría de las bacterias que utilizan alternativas del ciclo de Calvin son quimioautotróficas, también se ha demostrado que ciertas bacterias fotoautótrofas de azufre verde utilizan una vía alternativa de fijación de CO 2.
Ejercicio\(\PageIndex{3}\)
Describir las tres etapas del ciclo Calvino.
Conceptos clave y resumen
- Los heterótrofos dependen de los carbohidratos producidos por los autótrofos, muchos de los cuales son fotosintéticos, convirtiendo la energía solar en energía química.
- Diferentes organismos fotosintéticos utilizan diferentes mezclas de pigmentos fotosintéticos, que aumentan el rango de las longitudes de onda de luz que un organismo puede absorber.
- Los fotosistemas (PSI y PSII) contienen cada uno un complejo de recolección de luz, compuesto por múltiples proteínas y pigmentos asociados que absorben energía lumínica. Las reacciones de fotosíntesis dependientes de la luz convierten la energía solar en energía química, produciendo ATP y NADPH o NADH para almacenar temporalmente esta energía.
- En la fotosíntesis oxigénica, H 2 O sirve como donador de electrones para reemplazar el electrón central de reacción, y el oxígeno se forma como subproducto. En la fotosíntesis anoxigénica, otras moléculas reducidas como H 2 S o tiosulfato pueden ser utilizadas como donador de electrones; como tal, el oxígeno no se forma como subproducto.
- La fotofosforilación no cíclica se utiliza en la fotosíntesis oxigénica cuando se necesita tanto la producción de ATP como de NADPH. Si las necesidades de ATP de una célula superan sus necesidades de NADPH, entonces puede llevar a cabo fotofosforilación cíclica, produciendo solo ATP.
- Las reacciones de fotosíntesis independientes de la luz utilizan el ATP y el NADPH de las reacciones dependientes de la luz para fijar CO 2 en moléculas de azúcar orgánico.
Notas al pie
- 1 A. Dhingra et al. “Traducción Mejorada de un Gen Rbc S Expresado en Cloroplasto Restaura los Niveles de Subunidad Pequeña y la Fotosíntesis en Plantas Nucleares Rbc S antisentido” Actas de la Academia Nacional de Ciencias de los Estados Unidos de América 101 núm. 16 (2004) :6315—6320.