
10.1: Usando la Microbiología para Descubrir los Secretos de la Vida
- Page ID
- 54519

Objetivos de aprendizaje
- Describir el descubrimiento de ácidos nucleicos y nucleótidos
- Explicar los experimentos históricos que llevaron a la caracterización del ADN
- Describir cómo se han utilizado la microbiología y los microorganismos para descubrir la bioquímica de los genes
- Explicar cómo los científicos establecieron el vínculo entre el ADN y la herencia
Enfoque Clínico: Parte 1
Alex es un estudiante universitario de 22 años que estuvo de vacaciones en Puerta Vallarta, México, para las vacaciones de primavera. Desafortunadamente, dos días después de volar a su casa a Ohio, comenzó a experimentar calambres abdominales y diarrea acuosa extensa. Debido a su malestar, buscó atención médica en un gran hospital de Cincinnati cercano.
Ejercicio\(\PageIndex{1}\)
¿Qué tipos de infecciones u otras afecciones pueden ser responsables?
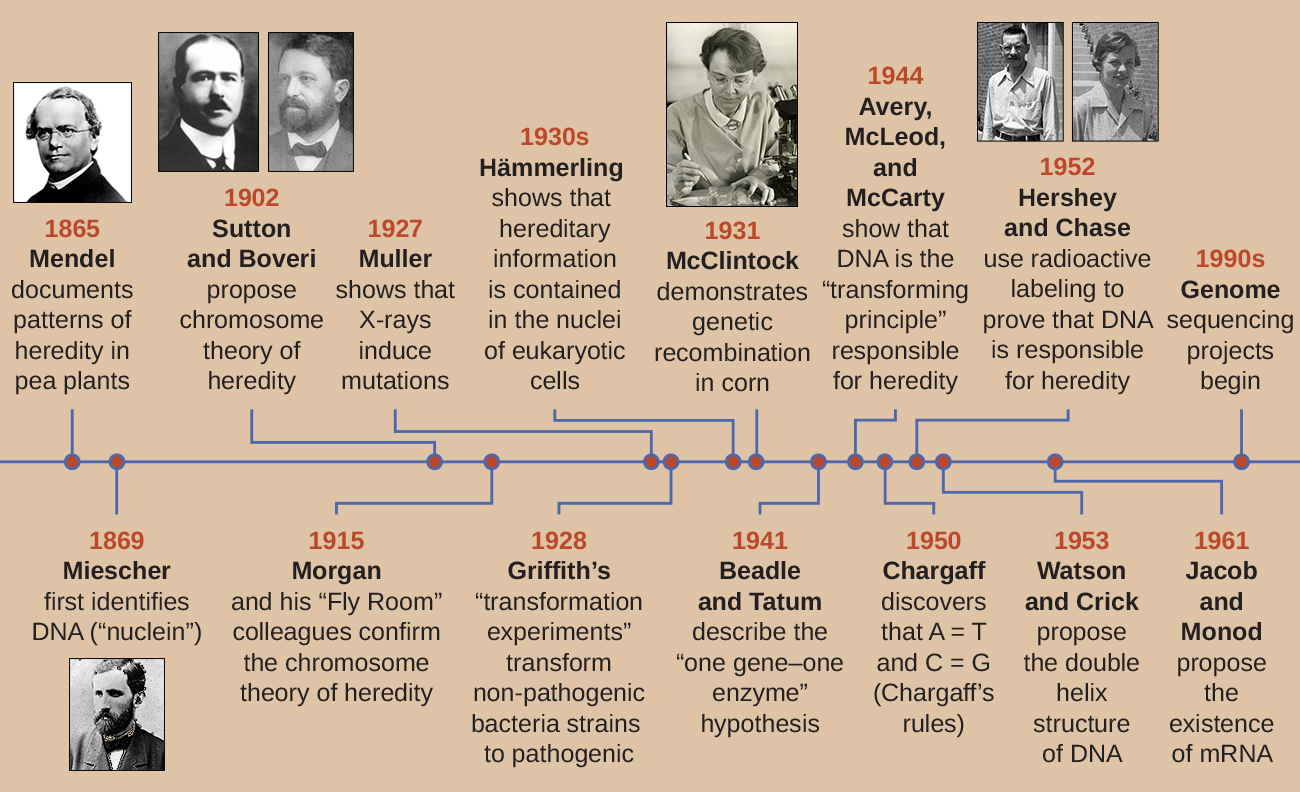
A principios del siglo XX, el ADN aún no fue reconocido como el material genético responsable de la herencia, el paso de rasgos de una generación a la siguiente. De hecho, gran parte de la investigación fue descartada hasta mediados del siglo XX. La comunidad científica creía, incorrectamente, que el proceso de herencia implicaba una mezcla de rasgos parentales que producían una apariencia física intermedia en la descendencia; este proceso hipotético parecía correcto por lo que hoy conocemos como variación continua, que resulta de la acción de muchos genes para determinar una característica particular, como la altura humana. La descendencia parece ser una “mezcla” de los rasgos de sus padres cuando observamos características que exhiben variación continua. La teoría mixta de la herencia aseveró que los rasgos parentales originales se perdieron o absorbieron por la mezcla en la descendencia, pero ahora sabemos que este no es el caso.
Dos líneas de investigación separadas, iniciadas a mediados y finales del siglo XIX, condujeron finalmente al descubrimiento y caracterización del ADN y los fundamentos de la genética, la ciencia de la herencia. Estas líneas de investigación comenzaron a converger en la década de 1920, y la investigación utilizando sistemas microbianos finalmente resultó en contribuciones significativas para dilucidar las bases moleculares de la genética.
Descubrimiento y Caracterización del ADN
La comprensión moderna del ADN ha evolucionado desde el descubrimiento del ácido nucleico hasta el desarrollo del modelo de doble hélice. En la década de 1860, Friedrich Miescher (1844—1895), médico de profesión, fue la primera persona en aislar químicos ricos en fósforo de los leucocitos (glóbulos blancos) del pus en vendajes usados de una clínica quirúrgica local. Llamó a estos químicos (que eventualmente se conocerían como ARN y ADN) “nucleina” porque estaban aislados de los núcleos de las células. Su alumno Richard Altmann (1852—1900) posteriormente lo calificó de “ácido nucleico” 20 años después cuando descubrió la naturaleza ácida de la nucleina. En las dos últimas décadas del siglo XIX, el bioquímico alemán Albrecht Kossel (1853-1927) aisló y caracterizó las cinco bases nucleotídicas diferentes que componen el ácido nucleico. Estos son adenina, guanina, citosina, timina (en ADN) y uracilo (en ARN). Kossell recibió el Premio Nobel de Fisiología o Medicina en 1910 por su trabajo sobre ácidos nucleicos y por su considerable trabajo sobre proteínas, incluido el descubrimiento de la histidina.
Fundamentos de la Genética
A pesar del descubrimiento del ADN a finales del siglo XIX, los científicos no hicieron la asociación con la herencia durante muchas décadas más. Para hacer esta conexión, los científicos, entre ellos varios microbiólogos, realizaron muchos experimentos con plantas, animales y bacterias.
Plantas de guisante de Mendel
Mientras Miescher aislaba y descubriría el ADN en la década de 1860, el monje y botánico austriaco Johann Gregor Mendel (1822—1884) experimentaba con arvejas de jardín, demostrando y documentando patrones básicos de herencia, ahora conocidas como leyes de Mendel.
En 1856, Mendel comenzó su investigación de una década sobre los patrones de herencia. Utilizó el guisante diploide de jardín, Pisum sativum, como su sistema modelo primario porque naturalmente se autofertiliza y es altamente endogámica, produciendo líneas de plantas de guisantes “de reproducción verdadera”, plantas que siempre producen descendencia que se parece al padre. Al experimentar con plantas de guisante de reproducción real, Mendel evitó la aparición de rasgos inesperados en la descendencia que podrían ocurrir si usaba plantas que no eran de reproducción real. Mendel realizó hibridaciones, las cuales implican el apareamiento de dos individuos de reproducción verdadera (generación P) que tienen rasgos diferentes, y examinó las características de su descendencia (primera generación filial, F 1) así como la descendencia de autofecundación de la generación F 1 (segunda generación filial, F 2) (Figura\(\PageIndex{1}\)).
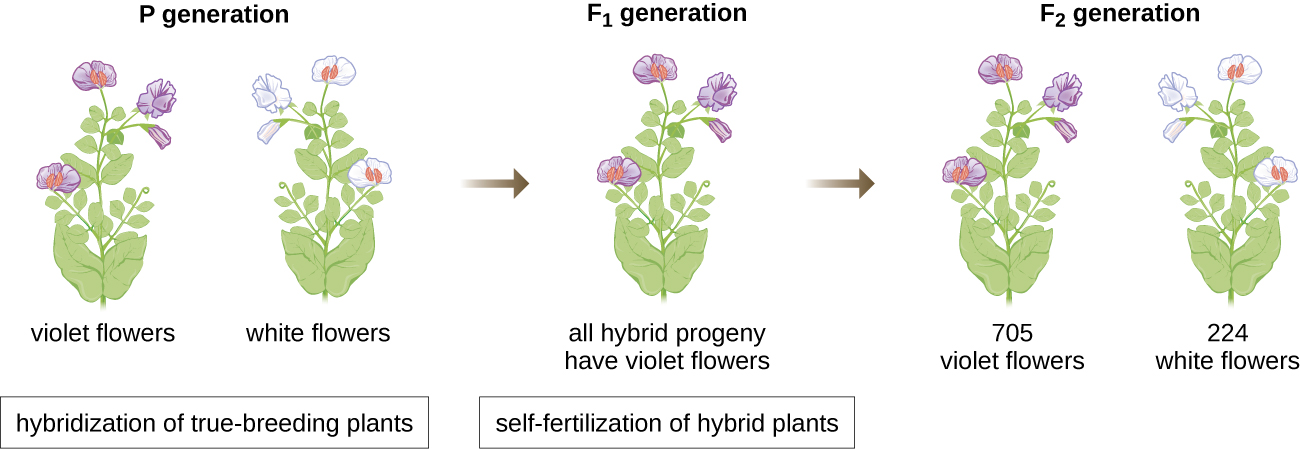
En 1865, Mendel presentó los resultados de sus experimentos con casi 30 mil plantas de guisantes a la sociedad local de historia natural. Demostró que los rasgos se transmiten fielmente de padres a hijos independientemente de otros rasgos. En 1866, publicó su obra, “Experimentos en hibridación de plantas”, 1 en las Actas de la Sociedad de Historia Natural de Brünn. El trabajo de Mendel pasó prácticamente desapercibido para la comunidad científica, que creyó, incorrectamente, en la teoría de la mezcla de rasgos en variación continua.
No fue reconocido por sus extraordinarias contribuciones científicas durante su vida. De hecho, no fue hasta 1900 que su obra fue redescubierta, reproducida y revitalizada por científicos a punto de descubrir las bases cromosómicas de la herencia.
La Teoría Cromosómica de la Herencia
Mendel realizó sus experimentos mucho antes de que los cromosomas fueran visualizados bajo un microscopio. Sin embargo, con la mejora de las técnicas microscópicas a finales del siglo XIX, los biólogos celulares pudieron teñir y visualizar estructuras subcelulares con tintes y observar sus acciones durante la meiosis. Fueron capaces de observar cromosomas replicándose, condensándose de una masa nuclear amorfa en distintos cuerpos en forma de X y migrando a polos celulares separados. La especulación de que los cromosomas podrían ser la clave para comprender la herencia llevó a varios científicos a examinar las publicaciones de Mendel y reevaluar su modelo en términos del comportamiento de los cromosomas durante la mitosis y la meiosis.
En 1902, Theodor Boveri (1862-1915) observó que en los erizos de mar, los componentes nucleares (cromosomas) determinaron el desarrollo embrionario adecuado. Ese mismo año, Walter Sutton (1877-1916) observó la separación de cromosomas en células hijas durante la meiosis. En conjunto, estas observaciones condujeron al desarrollo de la Teoría Cromosómica de la Herencia, que identificó a los cromosomas como el material genético responsable de la herencia mendeliana.
A pesar de las correlaciones convincentes entre el comportamiento de los cromosomas durante la meiosis y las observaciones de Mendel, la Teoría Cromosómica de la Herencia se propuso mucho antes de que existiera evidencia directa de que los rasgos se portaban en los cromosomas. Thomas Hunt Morgan (1866—1945) y sus colegas pasaron varios años realizando cruces con la mosca de la fruta, Drosophila melanogaster. Realizaron minuciosas observaciones microscópicas de cromosomas de mosca y correlacionaron estas observaciones con las características resultantes de la mosca. Su trabajo proporcionó la primera evidencia experimental para apoyar la Teoría Cromosómica de la Herencia a principios del siglo XX. En 1915, Morgan y sus colegas de “Fly Room” publicaron El mecanismo de la herencia mendeliana, que identificó a los cromosomas como las estructuras celulares responsables de la herencia. Por sus muchas contribuciones significativas a la genética, Morgan recibió el Premio Nobel de Fisiología o Medicina en 1933.
A finales de la década de 1920, Barbara McClintock (1902—1992) desarrolló técnicas de tinción cromosómica para visualizar y diferenciar entre los diferentes cromosomas del maíz (maíz). En las décadas de 1940 y 1950, identificó un evento de rotura en el cromosoma 9, al que llamó el locus de disociación (Ds). Ds podría cambiar de posición dentro del cromosoma. También identificó un locus activador (Ac). La rotura del cromosoma Ds podría ser activada por un elemento Ac (enzima transposasa). Al principio, el hallazgo de McClintock de estos genes saltadores, que ahora llamamos transposones, no fue aceptado por la comunidad científica. No fue hasta la década de 1960 y más tarde que los transposones fueron descubiertos en bacteriófagos, bacterias y Drosophila. Hoy en día, sabemos que los transposones son segmentos móviles de ADN que pueden moverse dentro del genoma de un organismo. Pueden regular la expresión génica, la expresión de proteínas y la virulencia (capacidad de causar enfermedades).
Microbios y virus en la investigación genética
Los microbiólogos también han jugado un papel crucial en nuestra comprensión de la genética. Los organismos experimentales como los guisantes de Mendel, las moscas de la fruta de Morgan y el maíz de McClintock ya se habían utilizado con éxito para allanar el camino para una comprensión de la genética. Sin embargo, los microbios y virus fueron (y siguen siendo) excelentes sistemas modelo para el estudio de la genética porque, a diferencia de los guisantes, las moscas de la fruta y el maíz, se propagan más fácilmente en el laboratorio, creciendo a altas densidades de población en una pequeña cantidad de espacio y en poco tiempo. Además, debido a su simplicidad estructural, los microbios y virus son más fácilmente manipulados genéticamente.
Afortunadamente, a pesar de diferencias significativas en tamaño, estructura, estrategias de reproducción y otras características biológicas, existe una unidad bioquímica entre todos los organismos; tienen en común las mismas moléculas subyacentes responsables de la herencia y del uso del material genético para dar a las células su variación características. En palabras del científico francés Jacques Monod, “Lo que es cierto para E. coli también es cierto para el elefante”, lo que significa que la bioquímica de la vida se ha mantenido a lo largo de la evolución y se comparte en todas las formas de vida, desde simples organismos unicelulares hasta organismos grandes y complejos. Esta continuidad bioquímica hace que los microbios sean excelentes modelos para usar en estudios genéticos.
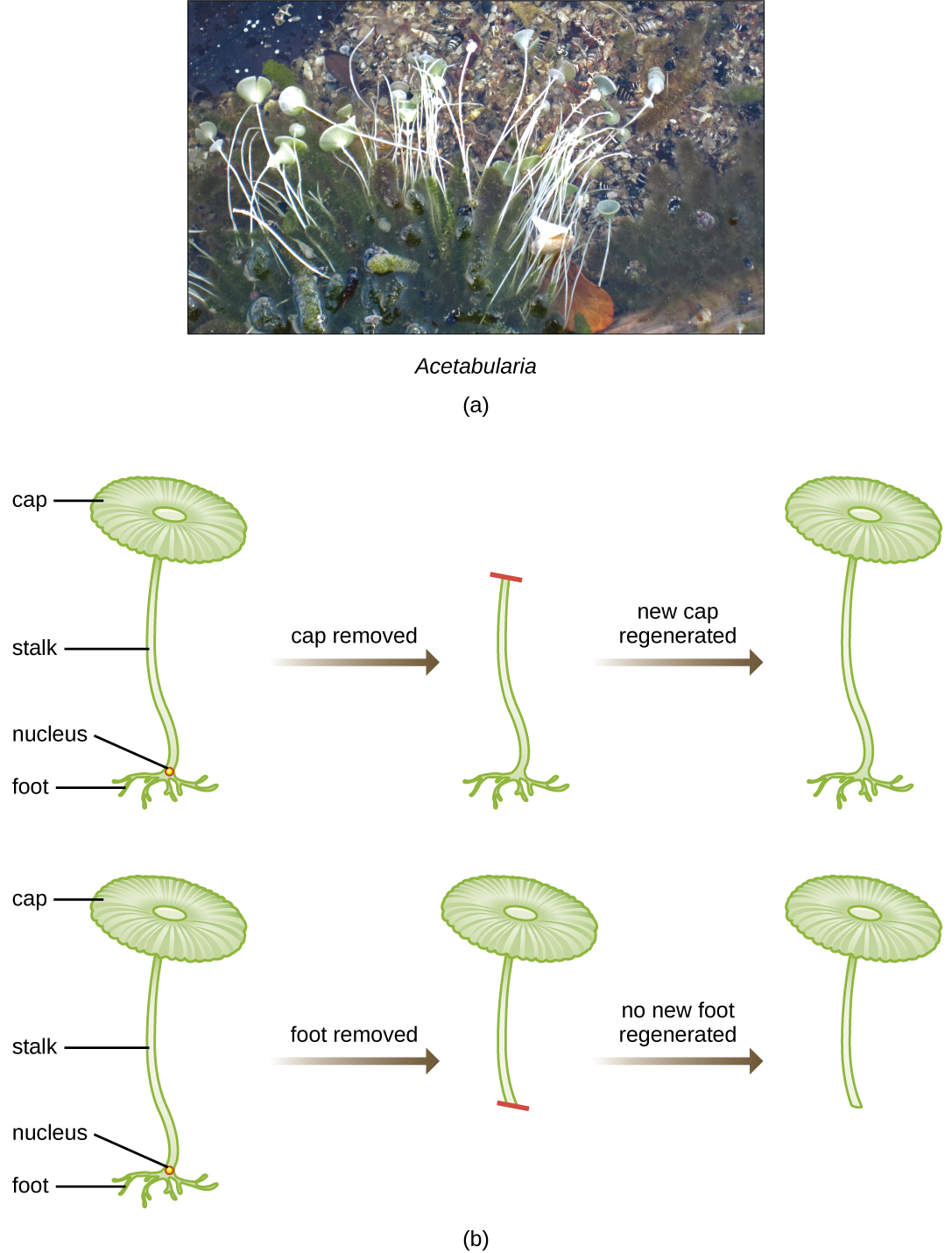
En un inteligente conjunto de experimentos en las décadas de 1930 y 1940, el científico alemán Joachim Hämmerling (1901—1980), utilizando la alga unicelular Acetabularia como modelo microbiano, estableció que la información genética en una célula eucariota se encuentra dentro del núcleo. Las acetabularias spp. son células de algas inusualmente grandes que crecen asimétricamente, formando un “pie” que contiene el núcleo, el cual se usa para la unión al sustrato; un tallo; y una tapa parecida a un paraguas, estructuras que se pueden ver fácilmente a simple vista. En un conjunto temprano de experimentos, Hämmerling quitó el capuchón o el pie de las células y observó si se regeneraban nuevas tapas o pies (Figura\(\PageIndex{2}\)). Encontró que cuando se quitó el pie de estas células, no crecieron nuevos pies; sin embargo, cuando se quitaron las tapas de las células, se regeneraron nuevas tapas. Esto sugiere que la información hereditaria se localizó en el pie que contenía núcleos de cada célula.
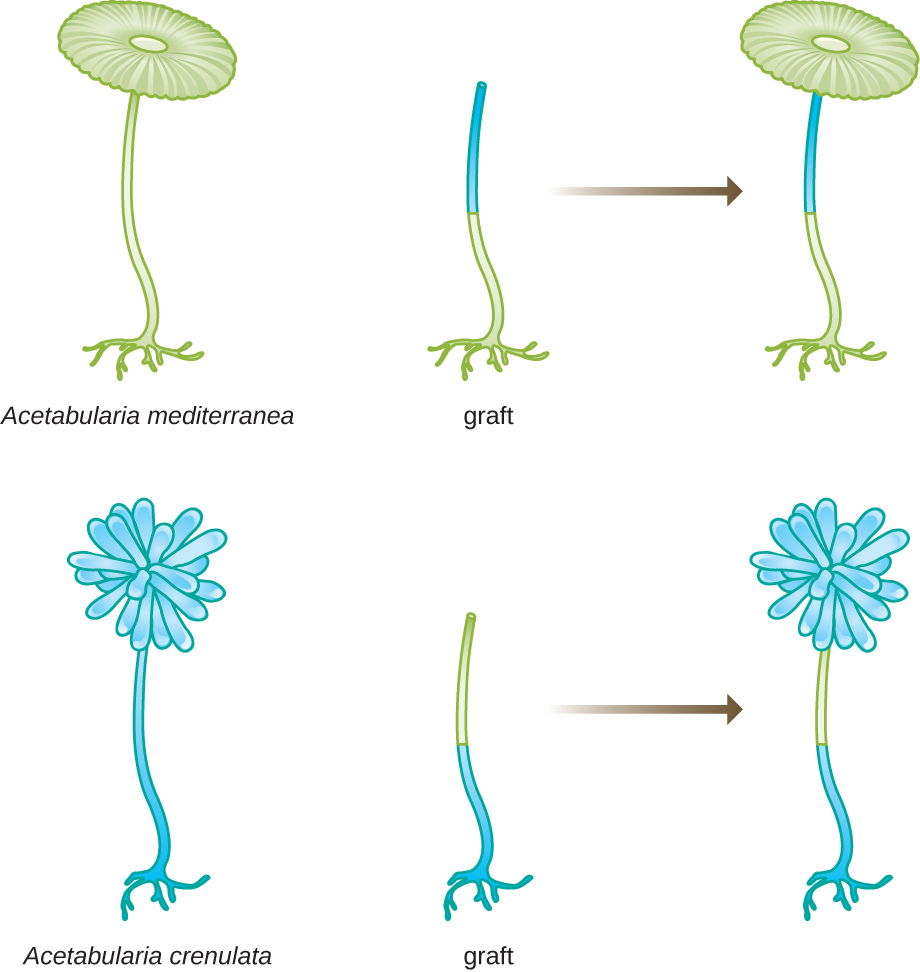
En otro conjunto de experimentos, Hämmerling utilizó dos especies de Acetabularia que tienen diferentes morfologías de caperuza, A. crenulata y A. mediterranea (Figura\(\PageIndex{3}\)). Cortó las tapas de ambos tipos de células y luego injertó el tallo de una A. crenulata en un pie de A. mediterranea, y viceversa. Con el tiempo, observó que la célula injertada con el pie de A. crenulata y el tallo de A. mediterranea desarrollaron una caperuza con la morfología de A. crenulata. Por el contrario, la célula injertada con el pie de A. mediterranea y el tallo de A. crenulata desarrollaron una caperuza con la morfología de A. mediterranea. Confirmó microscópicamente la presencia de núcleos en los pies de estas células y atribuyó el desarrollo de estas morfologías de caperuza al núcleo de cada célula injertada. Así, demostró experimentalmente que el núcleo era la ubicación del material genético que dictaba las propiedades de una célula.
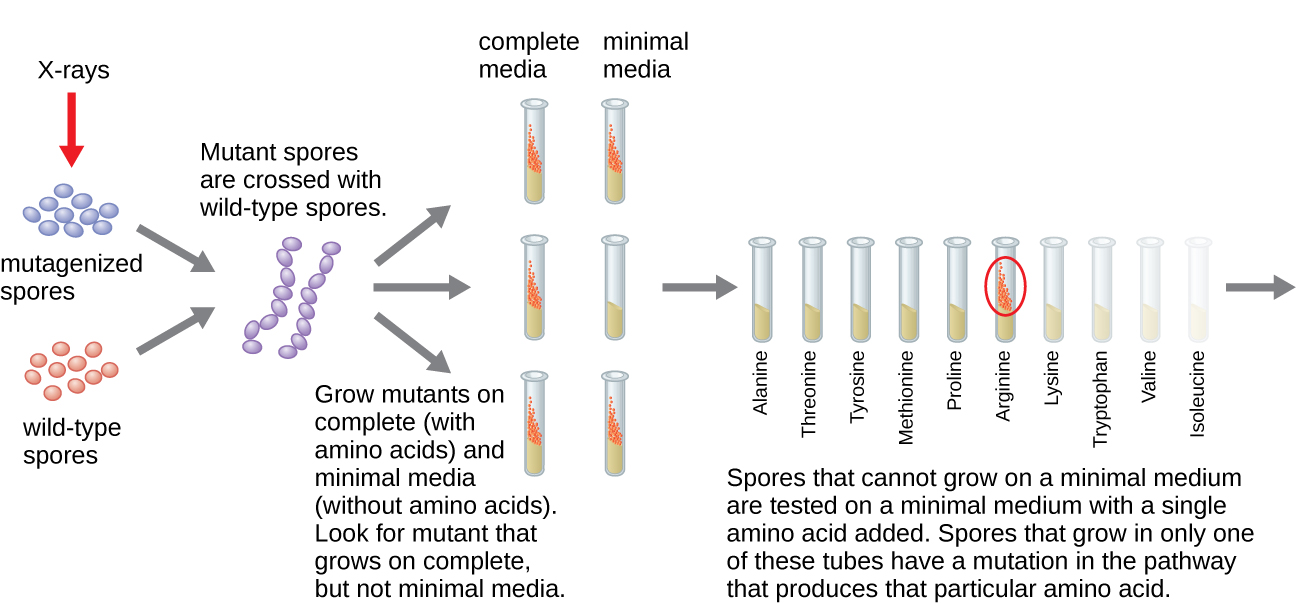
Otro modelo microbiano, el molde de pan rojo Neurospora crassa, fue utilizado por George Beadle y Edward Tatum para demostrar la relación entre los genes y las proteínas que codifican. Beadle había trabajado con moscas de la fruta en el laboratorio de Morgan pero las encontró demasiado complejas para realizar ciertos tipos de experimentos. N. crassa, por otro lado, es un organismo más sencillo y tiene la capacidad de crecer en un medio mínimo debido a que contiene vías enzimáticas que le permiten utilizar el medio para producir sus propias vitaminas y aminoácidos.
Beadle y Tatum irradiaron el moho con rayos X para inducir cambios en una secuencia de ácidos nucleicos, llamadas mutaciones. Se aparearon las esporas de moho irradiadas e intentaron cultivarlas tanto en un medio completo como en un medio mínimo. Buscaron mutantes que crecieran en un medio completo, complementados con vitaminas y aminoácidos, pero que no crecieran en el medio mínimo que carecía de estos suplementos. Dichos moldes contenían teóricamente mutaciones en los genes que codificaban las vías biosintéticas. Al encontrar tales mutantes, probaron sistemáticamente cada uno para determinar qué vitamina o aminoácido no pudo producir (Figura\(\PageIndex{4}\)) y publicaron este trabajo en 1941.
El trabajo posterior de Beadle, Tatum y colegas demostró que podían aislar diferentes clases de mutantes que requerían un suplemento particular, como el aminoácido arginina (Figura\(\PageIndex{5}\)). Con cierto conocimiento de la vía de biosíntesis de arginina, identificaron tres clases de mutantes de arginina complementando el medio mínimo con intermedios (citrulina u ornitina) en la vía. Los tres mutantes diferían en sus capacidades para crecer en cada uno de los medios, lo que llevó al grupo de científicos a proponer, en 1945, que cada tipo de mutante tenía un defecto en un gen diferente en la vía de biosíntesis de arginina. Esto condujo a la llamada hipótesis de un gen, una enzima, que sugería que cada gen codifica una enzima.
El conocimiento posterior sobre los procesos de transcripción y traducción llevó a los científicos a revisar esto a la hipótesis de “un gen, un polipéptido”. Aunque hay algunos genes que no codifican polipéptidos (sino que codifican para ARN de transferencia [ARNt] o ARN ribosómicos [ARNs], que discutiremos más adelante), la hipótesis de un gen, una enzima, es cierta en muchos casos, especialmente en microbios. El descubrimiento de Beadle y Tatum del vínculo entre los genes y las características correspondientes les valió el Premio Nobel de Fisiología y Medicina de 1958 y desde entonces se ha convertido en la base de la genética molecular moderna.
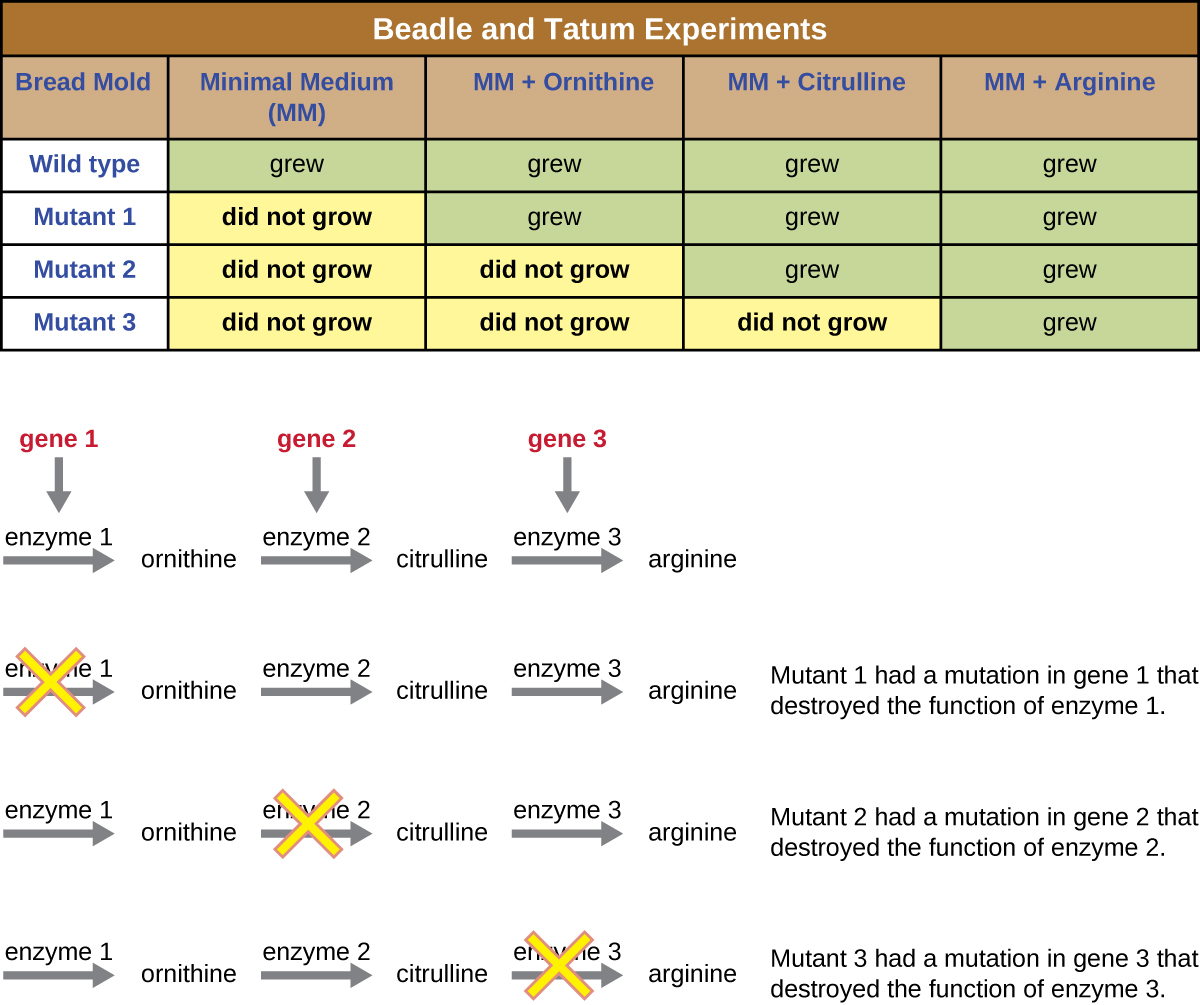
Para conocer más sobre los experimentos de Beadle y Tatum, visite este sitio web desde el Centro de Aprendizaje de ADN.
Ejercicio\(\PageIndex{2}\)
- ¿Qué organismo utilizaron Morgan y sus colegas para desarrollar la Teoría Cromosómica de la Herencia? ¿Qué rasgos rastrearon?
- ¿Qué demostró Hämmerling con sus experimentos con Acetabularia?
El ADN como la molécula responsable de la herencia
A principios del siglo XX ya se había realizado una gran cantidad de trabajo para caracterizar el ADN y establecer las bases de la genética, entre ellas atribuir herencia a los cromosomas que se encuentran dentro del núcleo. A pesar de toda esta investigación, no fue hasta bien entrado el siglo XX cuando estas líneas de investigación convergieron y los científicos comenzaron a considerar que el ADN podría ser el material genético que la descendencia heredó de sus padres. Se pensó que el ADN, que contenía solo cuatro nucleótidos diferentes, era estructuralmente demasiado simple para codificar información genética tan compleja. En cambio, se pensó que la proteína tenía la complejidad requerida para servir como información genética celular porque está compuesta por 20 aminoácidos diferentes que podrían combinarse en una gran variedad de combinaciones. Los microbiólogos desempeñaron un papel fundamental en la investigación que determinó que el ADN es la molécula responsable de la herencia.
Experimentos de transformación de Griffith
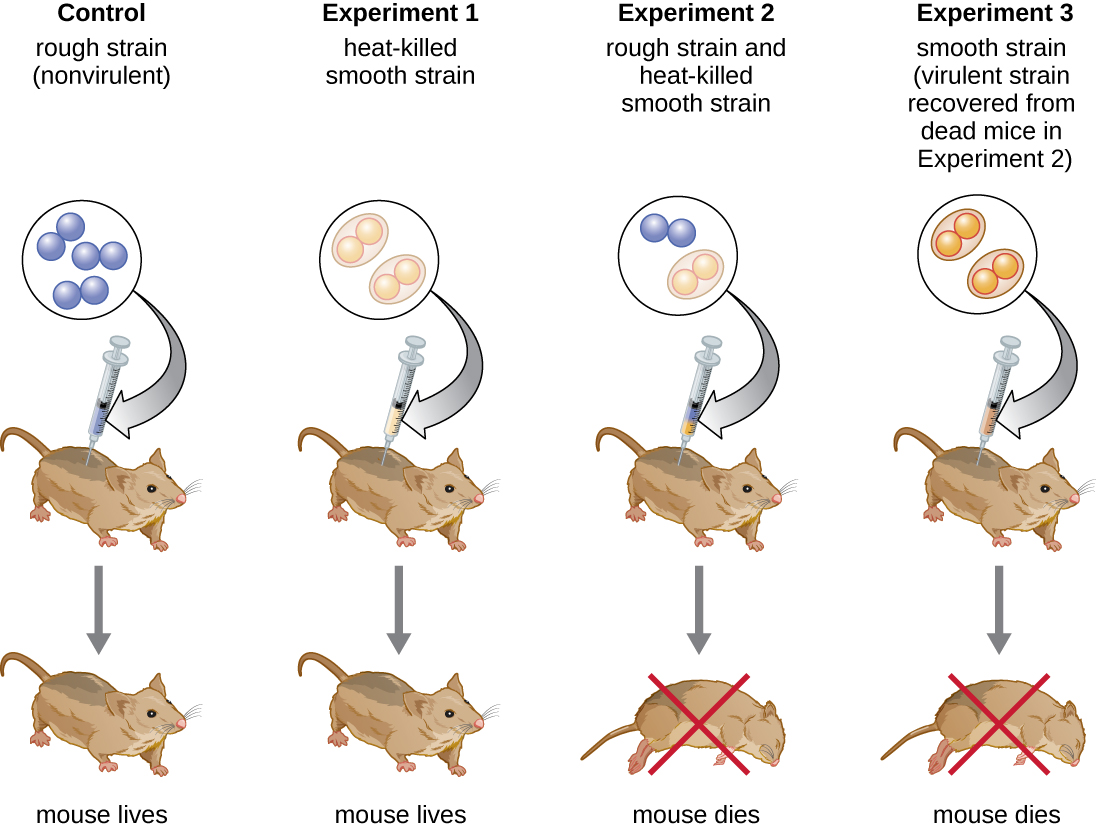
El bacteriólogo británico Frederick Griffith (1879—1941) fue quizás la primera persona en demostrar que la información hereditaria podía transferirse de una célula a otra “horizontalmente” (entre miembros de la misma generación), en lugar de “verticalmente” (de padre a descendencia). En 1928, reportó la primera demostración de transformación bacteriana, proceso en el que el ADN externo es absorbido por una célula, cambiando así sus características. 3 Estaba trabajando con dos cepas de Streptococcus pneumoniae, una bacteria causante de neumonía: una cepa rugosa (R) y una cepa lisa (S). La cepa R es no patógena y carece de una cápsula en su superficie externa; como resultado, las colonias de la cepa R aparecen ásperas cuando se cultivan en placas. La cepa S es patógena y tiene una cápsula fuera de su pared celular, lo que le permite escapar de la fagocitosis por parte del sistema inmune del huésped. Las cápsulas hacen que las colonias de la cepa S aparezcan lisas cuando se cultivan en placas.
En una serie de experimentos, Griffith analizó los efectos de las cepas R vivas, S vivas y S muertas por calor de S. pneumoniae en ratones vivos (Figura\(\PageIndex{6}\)). Cuando se inyectó a los ratones la cepa S viva, los ratones murieron. Cuando inyectó a los ratones la cepa R viva o la cepa S destruida por calor, los ratones sobrevivieron. Pero cuando inyectó a los ratones una mezcla de cepa R viva y cepa S destruida por calor, los ratones murieron. Al aislar las bacterias vivas del ratón muerto, solo recuperó la cepa S de bacterias. Cuando luego inyectó esta cepa S aislada en ratones frescos, los ratones murieron. Griffith concluyó que algo había pasado de la cepa S destruida por calor a la cepa R viva y la “transformó” en la cepa S patógena; llamó a esto el “principio transformador”. Estos experimentos se conocen ahora como experimentos de transformación de Griffith.
En 1944, Oswald Avery, Colin MacLeod y Maclyn McCarty estaban interesados en explorar más a fondo el principio transformador de Griffith. Aislaron la cepa S de ratones muertos infectados, la mataron por calor e inactivaron varios componentes del extracto de S, realizando un estudio de eliminación sistemática (Figura\(\PageIndex{7}\)). Utilizaron enzimas que degradaban específicamente proteínas, ARN y ADN y mezclaban el extracto S con cada una de estas enzimas individuales. Luego, probaron la capacidad resultante de cada combinación de extracto/enzima para transformar la cepa R, como se observó por el crecimiento difuso de la cepa S en medios de cultivo y se confirmó visualmente por el crecimiento en placas. Encontraron que cuando el ADN se degradaba, la mezcla resultante ya no era capaz de transformar la bacteria de la cepa R, mientras que ningún otro tratamiento enzimático pudo evitar la transformación. Esto los llevó a concluir que el ADN era el principio transformador. A pesar de sus resultados, muchos científicos no aceptaron su conclusión, sino que creyeron que había contaminantes proteicos dentro de sus extractos.
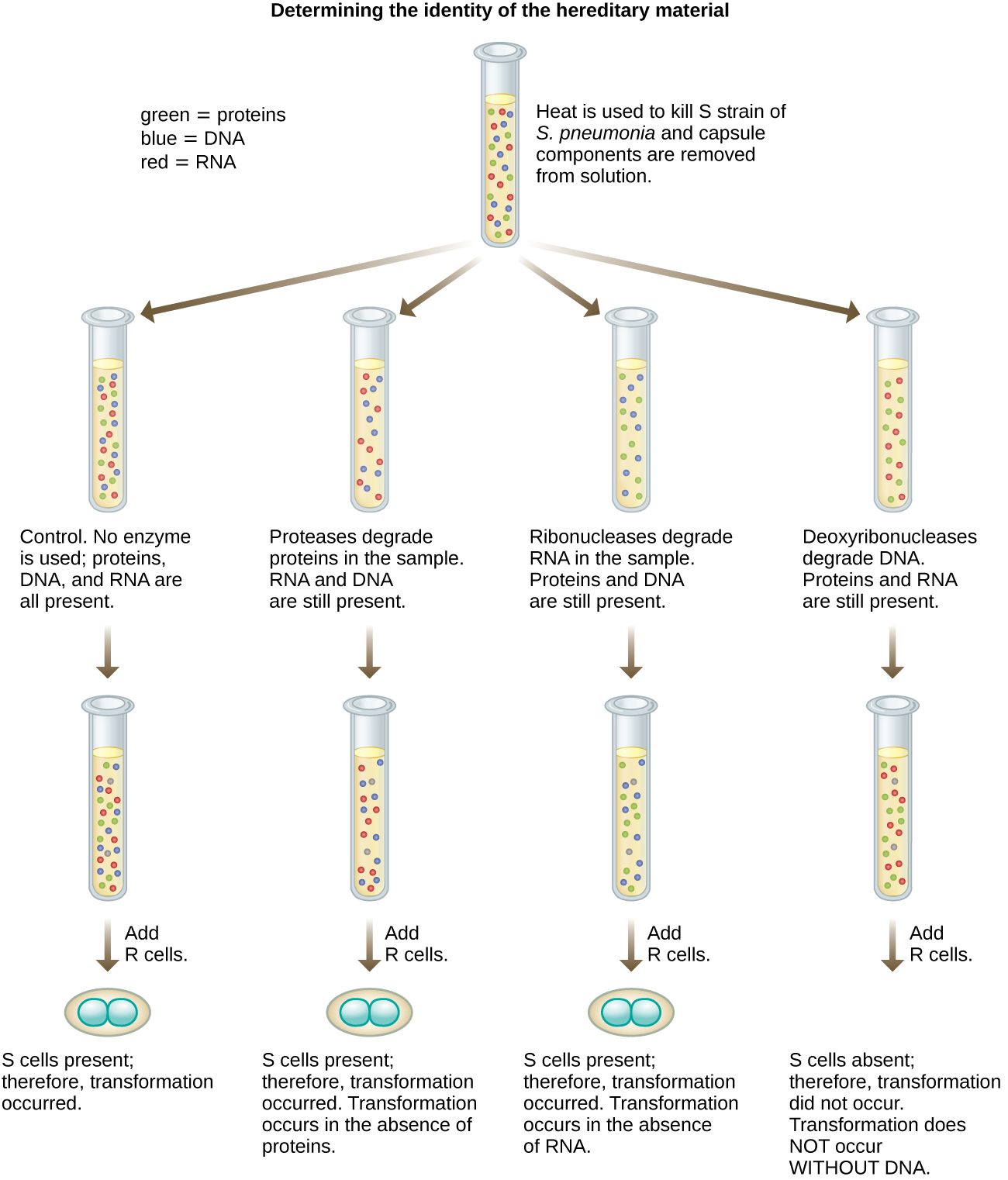
Ejercicio\(\PageIndex{3}\)
¿Cómo demostraron los experimentos de Avery, MacLeod y McCarty que el ADN era el principio transformador descrito por primera vez por Griffith?
Prueba de ADN como material genético de Hershey y Chase
Alfred Hershey y Martha Chase realizaron sus propios experimentos en 1952 y pudieron proporcionar evidencia confirmatoria de que el ADN, no la proteína, era el material genético (Figura\(\PageIndex{8}\)). 4 Hershey y Chase estudiaban un bacteriófago, un virus que infecta bacterias. Los virus suelen tener una estructura simple: una cubierta proteica, llamada cápside, y un núcleo de ácido nucleico que contiene el material genético, ya sea ADN o ARN (ver Virus). El bacteriófago particular que estaban estudiando fue el bacteriófago T2, el cual infecta células de E. coli. Como hoy sabemos, T2 se adhiere a la superficie de la célula bacteriana y luego inyecta sus ácidos nucleicos dentro de la célula. El ADN del fago hace múltiples copias de sí mismo usando la maquinaria del huésped, y eventualmente la célula huésped estalla, liberando una gran cantidad de bacteriófagos.
Hershey y Chase marcaron la cubierta proteica en un lote de fagos usando azufre radiactivo, 35 S, porque el azufre se encuentra en los aminoácidos metionina y cisteína pero no en los ácidos nucleicos. Marcaron el ADN en otro lote usando fósforo radiactivo, 32 P, debido a que el fósforo se encuentra en el ADN y el ARN pero no típicamente en la proteína.
Cada lote de fagos se dejó infectar las células por separado. Después de la infección, Hershey y Chase colocaron cada suspensión bacteriana de fago en una licuadora, que separó las capas de fago de la célula hospedadora y centrifugaron la suspensión resultante en una centrífuga. Las células bacterianas más pesadas se asentaron y formaron un sedimento, mientras que las partículas de fago más ligeras permanecieron en el sobrenadante. En el tubo con la proteína marcada, la radiactividad permaneció sólo en el sobrenadante. En el tubo con el ADN marcado, la radiactividad se detectó únicamente en las células bacterianas. Hershey y Chase concluyeron que fue el ADN del fago el que se inyectó en la célula el que portaba la información para producir más partículas de fago, demostrando así que el ADN, no las proteínas, era la fuente del material genético. Como resultado de su trabajo, la comunidad científica aceptó más ampliamente el ADN como la molécula responsable de la herencia.
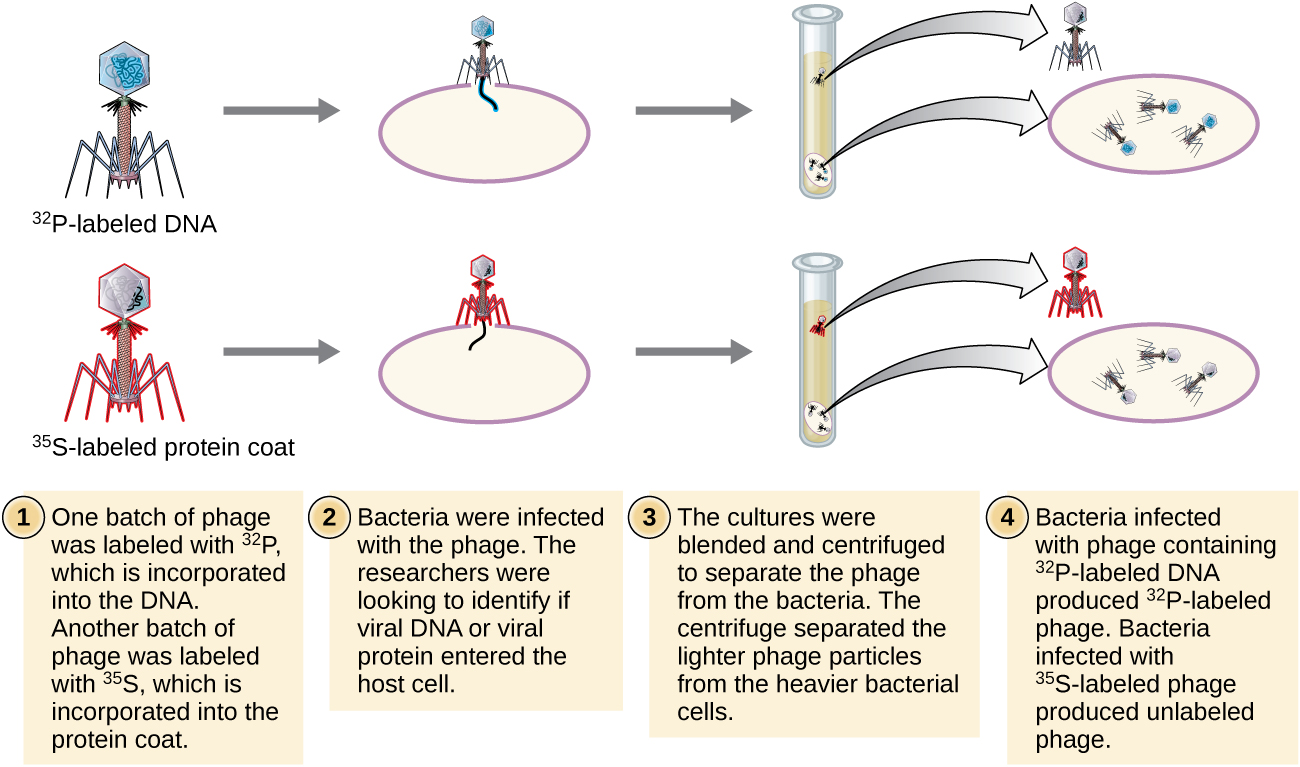
Para cuando Hershey y Chase publicaron su experimento a principios de la década de 1950, los microbiólogos y otros científicos habían estado investigando la herencia durante más de 80 años. Basándose en la investigación de los demás durante ese tiempo culminó con el acuerdo general de que el ADN era el material genético responsable de la herencia (Figura\(\PageIndex{9}\)). Este conocimiento sentó las bases para la era de la biología molecular por venir y los avances significativos en biotecnología y biología de sistemas que estamos viviendo hoy en día.
Para conocer más sobre los experimentos involucrados en la historia de la genética y el descubrimiento del ADN como material genético de las células, visite este sitio web desde el Centro de Aprendizaje de ADN.
Ejercicio\(\PageIndex{4}\)
¿Cómo usaron Hershey y Chase los microbios para demostrar que el ADN es material genético?
Conceptos clave y resumen
- El ADN fue descubierto y caracterizado mucho antes de que se entendiera su papel en la herencia. Los microbiólogos desempeñaron un papel significativo en demostrar que el ADN es la información hereditaria que se encuentra dentro de las
- En las décadas de 1850 y 1860, Gregor Mendel experimentó con guisantes de cultivo real para demostrar la heredabilidad de rasgos específicos observables.
- En 1869, Friedrich Miescher aisló y purificó un compuesto rico en fósforo de los núcleos de los glóbulos blancos; nombró al compuesto nucleina. El estudiante de Miescher, Richard Altmann, descubrió su naturaleza ácida, renombrándolo como ácido nucleico. Albrecht Kossell caracterizó las bases nucleotídicas encontradas dentro de los ácidos nucleicos.
- Aunque Walter Sutton y Theodor Boveri propusieron la Teoría Cromosómica de la Herencia en 1902, no se demostró científicamente hasta la publicación en 1915 de la obra de Thomas Hunt Morgan y sus colegas.
- Utilizando Acetabularia, una gran célula de algas, como su sistema modelo, Joachim Hämmerling demostró en las décadas de 1930 y 1940 que el núcleo era la ubicación de información hereditaria en estas células.
- En la década de 1940, George Beadle y Edward Tatum utilizaron el molde Neurospora crassa para demostrar que la producción de cada proteína estaba bajo el control de un solo gen, demostrando la hipótesis de “un gen—una enzima”.
- En 1928, Frederick Griffith demostró que las bacterias encapsuladas muertas podían pasar información genética a bacterias vivas no encapsuladas y transformarlas en cepas dañinas. En 1944, Oswald Avery, Colin McLeod y Maclyn McCarty identificaron el compuesto como ADN.
- La naturaleza del ADN como molécula que almacena información genética se demostró inequívocamente en el experimento de Alfred Hershey y Martha Chase publicado en 1952. El ADN marcado de virus bacterianos ingresó e infectó a las células bacterianas, dando lugar a más partículas virales. Las capas proteicas etiquetadas no participaron en la transmisión de información genética.
Notas al pie
- 1 J.G. Mendel. “Versuche über Pflanzenhybriden.” Verhandlungen des naturforschenden Vereines en Brünn, Bd. Abhandlungen 4 (1865) :3—7. (Para traducción al inglés, consulte http://www.mendelweb.org/Mendel.plain.html)
- 2 G.W. Beadle, E.L. Tatum. “Control Genético de Reacciones Bioquímicas en Neurospora”. Actas de la Academia Nacional de Ciencias 27 núm. 11 (1941) :499—506.
- 3 F. Griffith. “La Importancia de los Tipos Neumocócicos”. Revista de Higiene 27 núm. 2 (1928) :8—159.
- 4 A.D. Hershey, M. Chase. “Funciones Independientes de la Proteína Viral y el Ácido Nucleico en el Crecimiento del Bacteriófago”. Revista de Fisiología General 36 núm. 1 (1952) :39—56.