
11.6: Cómo los procariotas asexuales logran la diversidad genética
- Page ID
- 54599

Objetivos de aprendizaje
- Comparar los procesos de transformación, transducción y conjugación
- Explicar cómo la transferencia de genes asexuales da como resultado la diversidad genética procariota
- Explicar la estructura y consecuencias para la diversidad genética bacteriana de los transposones
Normalmente, cuando consideramos la transferencia genética, pensamos en la transferencia vertical de genes, la transmisión de información genética de generación en generación. La transferencia vertical de genes es, con mucho, el principal modo de transmisión de información genética en todas las células. En los organismos de reproducción sexual, los eventos de cruce y el surtido independiente de cromosomas individuales durante la meiosis contribuyen a la diversidad genética en la población. La diversidad genética también se introduce durante la reproducción sexual, cuando se combina la información genética de dos padres, cada uno con diferentes complementos de información genética, produciendo nuevas combinaciones de genotipos parentales en la descendencia diploide. La ocurrencia de mutaciones también contribuye a la diversidad genética en una población. La diversidad genética de la descendencia es útil en entornos cambiantes o inconsistentes y puede ser una razón para el éxito evolutivo de la reproducción sexual.
Cuando procariotas y eucariotas se reproducen asexualmente, transfieren una copia casi idéntica de su material genético a su descendencia a través de la transferencia vertical de genes. Aunque la reproducción asexual produce más descendencia con mayor rapidez, se pierde cualquier beneficio de la diversidad entre esas crías. ¿Cómo, entonces, los organismos cuyo modo reproductivo dominante es asexual crean diversidad genética? En procariotas, la transferencia génica horizontal (HGT), la introducción de material genético de un organismo a otro organismo dentro de una misma generación, es una forma importante de introducir la diversidad genética. El HGT permite que incluso especies distantemente relacionadas compartan genes, influyendo en sus fenotipos. Se piensa que el HGT es más prevalente en procariotas pero que solo una pequeña fracción del genoma procariota puede ser transferida por este tipo de transferencia en cualquier momento. A medida que el fenómeno se investiga más a fondo, puede revelarse que es aún más común. Muchos científicos creen que el HGT y la mutación son fuentes significativas de variación genética, la materia prima para el proceso de selección natural, en procariotas. Aunque el HGT es más común entre los organismos evolutivamente relacionados, puede ocurrir entre dos especies que conviven en una comunidad natural.
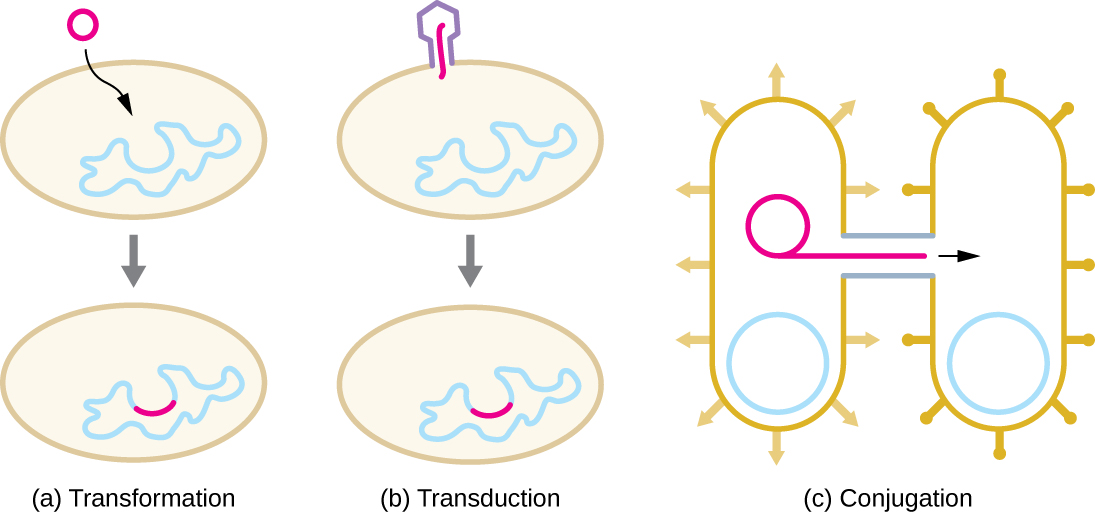
Se sabe que el HGT en procariotas ocurre por los tres mecanismos primarios que se ilustran en la Figura\(\PageIndex{1}\):
- Transformación: el ADN desnudo es tomado del ambiente
- Transducción: los genes se transfieren entre células en un virus (ver El ciclo de vida viral)
- Conjugación: uso de un tubo hueco llamado pilus de conjugación para transferir genes entre células
Ejercicio\(\PageIndex{1}\)
- ¿Cuáles son tres formas en que la reproducción sexual introduce la variación genética en la descendencia
- ¿Qué es un beneficio de la reproducción asexual?
- ¿Cuáles son los tres mecanismos de transferencia horizontal de genes en procariotas?
Transformación
Frederick Griffith fue el primero en demostrar el proceso de transformación. En 1928, demostró que la bacteria Streptococcus pneumoniae, no patógena y viva, podría transformarse en bacterias patógenas a través de la exposición a una cepa patógena destruida por calor. Concluyó que algún tipo de agente, al que llamó el “principio transformador”, había pasado de las bacterias patógenas muertas a las bacterias vivas, no patógenas. En 1944, Oswald Avery (1877—1955), Colin MacLeod (1909—1972) y Maclyn McCarty (1911—2005) demostraron que el principio transformador era el ADN (ver Usando la microbiología para descubrir los secretos de la vida).
En transformación, el procariota retoma ADN desnudo que se encuentra en su entorno y que se deriva de otras células que se han lisado al morir y han liberado su contenido, incluido su genoma, al ambiente. Muchas bacterias son naturalmente competentes, lo que significa que se unen activamente al ADN ambiental, lo transportan a través de sus envolturas celulares hacia su citoplasma y lo convierten en monocatenario. Por lo general, el ADN extraño bicatenario dentro de las células es destruido por nucleasas como defensa contra la infección viral. Sin embargo, estas nucleasas suelen ser ineficaces contra el ADN monocatenario, por lo que este ADN monocatenario dentro de la célula tiene la oportunidad de recombinarse en el genoma bacteriano. Una molécula de ADN que contiene fragmentos de ADN de diferentes organismos se llama ADN recombinante. (El ADN recombinante se discutirá con más detalle en Microbios y las Herramientas de Ingeniería Genética.) Si la bacteria incorpora el nuevo ADN en su propio genoma a través de la recombinación, la célula bacteriana puede obtener nuevas propiedades fenotípicas. Por ejemplo, si una bacteria no patógena toma ADN para un gen de toxina de un patógeno y luego lo incorpora a su cromosoma, también puede volverse patógena. El ADN plasmídico también puede ser captado por bacterias competentes y conferir nuevas propiedades a la célula. En general, la transformación en la naturaleza es un proceso relativamente ineficiente porque los niveles ambientales de ADN son bajos debido a la actividad de nucleasas que también se liberan durante la lisis celular. Adicionalmente, la recombinación genética es ineficiente para incorporar nuevas secuencias de ADN en el genoma.
En la naturaleza, la transformación bacteriana es un mecanismo importante para la adquisición de elementos genéticos que codifican factores de virulencia y resistencia a antibióticos. Se ha demostrado que los genes que codifican resistencia a compuestos antimicrobianos están muy extendidos en la naturaleza, incluso en entornos no influenciados por humanos. Estos genes, que permiten que los microbios que viven en comunidades mixtas compitan por recursos limitados, pueden ser transferidos dentro de una población por transformación, así como por los otros procesos de HGT. En el laboratorio, podemos explotar el proceso natural de transformación bacteriana para la ingeniería genética para elaborar una amplia variedad de medicamentos, como se discute en Microbios y las Herramientas de Ingeniería Genética.
Ejercicio\(\PageIndex{2}\)
¿Por qué una célula bacteriana produce ADN ambiental introducido en la célula en una forma monocatenaria?
Transducción
Los virus que infectan bacterias (bacteriófagos) también pueden mover trozos cortos de ADN cromosómico de una bacteria a otra en un proceso llamado transducción (ver Figura 6.2.3). Recordemos que en la transducción generalizada, cualquier fragmento de ADN cromosómico puede transferirse a una nueva célula hospedadora mediante el empaquetamiento accidental de ADN cromosómico en una cabeza de fago durante el ensamblaje del fago. Por el contrario, la transducción especializada resulta de la escisión imprecisa de un profago lisogénico del cromosoma bacteriano de tal manera que lleva consigo una parte del cromosoma bacteriano desde ambos lados del sitio de integración del fago a una nueva célula hospedadora. Como resultado, el anfitrión puede adquirir nuevas propiedades. Este proceso se llama conversión lisogénica. De importancia médica, un fago lisogénico puede llevar consigo un gen de virulencia a su nuevo huésped. Una vez insertado en el cromosoma del nuevo huésped, el nuevo hospedador puede ganar patogenicidad. Varias bacterias patógenas, entre ellas Corynebacterium diphtheriae (el agente causante de la difteria) y Clostridium botulinum (el agente causante del botulismo), son virulentas por la introducción de genes codificantes de toxinas por bacteriófagos lisogénicos, afirmando la clínica relevancia de la transducción en el intercambio de genes involucrados en enfermedades infecciosas. Las arqueas tienen sus propios virus que translocan material genético de un individuo a otro.
Ejercicio\(\PageIndex{3}\)
- ¿Cuál es el agente de transducción de células procariotas?
- En transducción especializada, ¿de dónde viene la pieza transductora de ADN?
Las consecuencias clínicas de la transducción
Paul, un trabajador de socorro de 23 años de Atlanta, viajó a Haití en 2011 para brindar ayuda tras el terremoto de 2010. Después de trabajar allí durante varias semanas, de repente comenzó a experimentar angustia abdominal, incluyendo calambres severos, náuseas, vómitos y diarrea acuosa. También comenzó a experimentar intensos calambres musculares. En una clínica local, el médico sospechó que los síntomas de Paul fueron causados por el cólera porque había habido un brote de cólera tras el sismo. Debido a que el cólera se transmite por vía fecal-oral, las brechas en la infraestructura de saneamiento, como suelen ocurrir después de desastres naturales, pueden precipitar brotes. El médico confirmó el diagnóstico presuntivo mediante una prueba de varilla reactiva del cólera. Luego le recetó a Paul una sola dosis de doxiciclina, así como sales de rehidratación oral, instruyéndole a beber cantidades significativas de agua limpia.
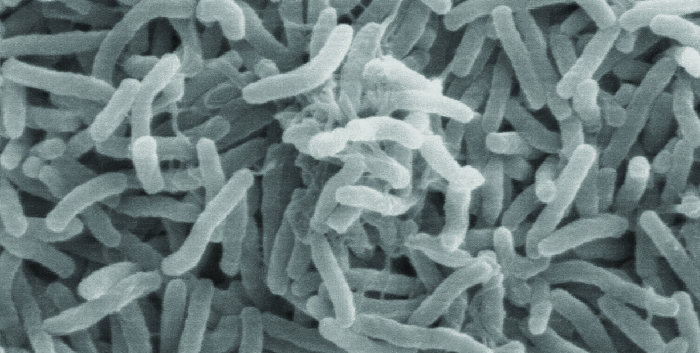
El cólera es causado por la varilla curva gramnegativa Vibrio cholerae (Figura\(\PageIndex{2}\)). Sus síntomas son en gran parte el resultado de la producción de la toxina del cólera (CT), que finalmente activa un transportador de cloruro para bombear iones cloruro fuera de las células epiteliales hacia el lumen intestinal. Luego, el agua sigue a los iones cloruro, provocando la prolífica diarrea acuosa característica del cólera. El gen que codifica la toxina del cólera se incorpora al cromosoma bacteriano de V. cólera a través de la infección de la bacteria con el fago CTX filamentoso lisogénico, que porta el gen CT y lo introduce en el cromosoma en la integración del profago. Así, las cepas patógenas de V. cholerae son el resultado de la transferencia horizontal de genes por transducción especializada.
Ejercicio\(\PageIndex{4}\)
- ¿Por qué son más comunes los brotes de cólera como consecuencia de un desastre natural?
- ¿Por qué los calambres musculares son un síntoma común del cólera? ¿Por qué es tan importante el tratamiento con sales de rehidratación oral para el tratamiento del cólera?
- En zonas afectadas por el cólera, ¿cuáles son algunas estrategias que las personas podrían utilizar para prevenir la transmisión de enfermedades?
Conjugación
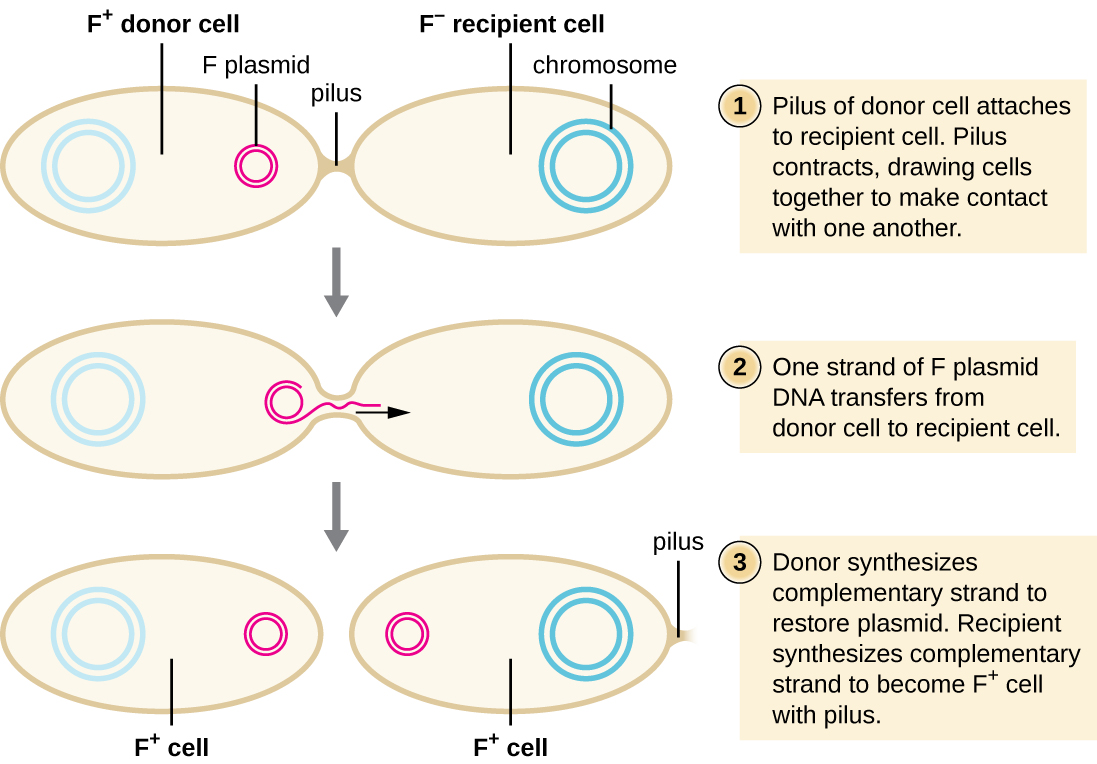
En conjugación, el ADN se transfiere directamente de un procariota a otro por medio de un pilus de conjugación, que pone los organismos en contacto entre sí. En E. coli, los genes que codifican la capacidad de conjugación se localizan en un plásmido bacteriano llamado plásmido F, también conocido como factor de fertilidad, y el pilus de conjugación se llama pilus F. Los genes del plásmido F codifican tanto las proteínas que componen el pilus F como las involucradas en la replicación en círculo rodante del plásmido. Las células que contienen el plásmido F, capaces de formar un pilus F, se denominan células F + o células donantes, y las que carecen de un plásmido F se denominan células F − o células receptoras.
Conjugación del plásmido F
Durante la conjugación típica en E. coli, el pilus F de una célula F + entra en contacto con una célula F y se retrae, poniendo en contacto las dos envolturas celulares (Figura\(\PageIndex{3}\)). Luego se forma un puente citoplásmico entre las dos células en el sitio del pilus de conjugación. A medida que la replicación en círculo rodante del plásmido F ocurre en la célula F +, una copia monocatenaria del plásmido F se transfiere a través del puente citoplásmico a la célula F −, que luego sintetiza la cadena complementaria, convirtiéndola en bicatenaria. La célula F − se convierte ahora en una célula F + capaz de hacer su propio pilus de conjugación. Eventualmente, en una población bacteriana mixta que contiene tanto células F + como F −, todas las células se convertirán en células F +. Los genes en el plásmido F de E. coli también codifican proteínas que previenen la conjugación entre células F +.
Conjugación de células F' y Hfr
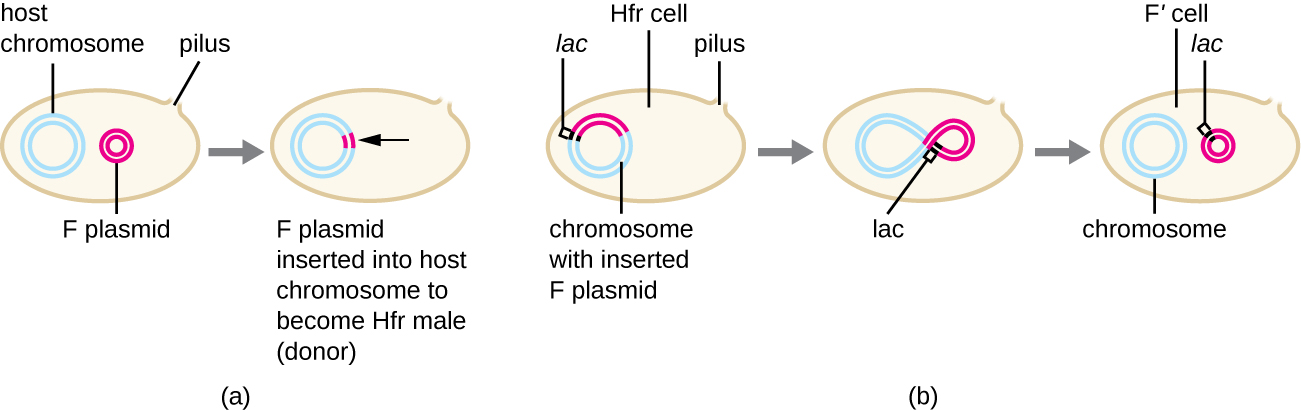
Aunque la conjugación típica en E. coli da como resultado la transferencia del ADN del plásmido F solamente, la conjugación también puede transferir ADN cromosómico. Esto se debe a que el plásmido F ocasionalmente se integra en el cromosoma bacteriano a través de la recombinación entre el plásmido y el cromosoma, formando una célula Hfr (Figura\(\PageIndex{4}\)). “" "Hfr "” se refiere a la alta frecuencia de recombinación observada cuando las células F − receptoras reciben información genética de células Hfr a través de la conjugación.” Similar a la escisión imprecisa de un profago durante la transducción especializada, el plásmido F integrado también se puede extirpar imprecisamente del cromosoma, produciendo un plásmido F' que lleva consigo algún ADN cromosómico adyacente al sitio de integración. En la conjugación, este ADN se introduce en la célula receptora y puede mantenerse como parte del plásmido F' o recombinarse en el cromosoma bacteriano de la célula receptora.
Las células Hfr también pueden tratar el cromosoma bacteriano como un enorme plásmido F e intentar transferir una copia del mismo a una célula F − receptora. Debido a que el cromosoma bacteriano es tan grande, la transferencia de todo el cromosoma lleva mucho tiempo (Figura\(\PageIndex{5}\)). Sin embargo, el contacto entre células bacterianas durante la conjugación es transitorio, por lo que es inusual que se transfiera todo el cromosoma. El ADN cromosómico del huésped cerca del sitio de integración del plásmido F, desplazado por el proceso unidireccional de replicación en círculo rodante, es más probable que se transfiera y recombinara en el cromosoma de una célula receptora que los genes hospedadores más alejados. Así, la ubicación relativa de los genes bacterianos en el genoma de la célula Hfr puede mapearse en función de cuándo se transfieren a través de la conjugación. Como resultado, antes de la edad de la secuenciación generalizada del genoma bacteriano, las distancias en los mapas genómicos procariotas a menudo se midieron en minutos.
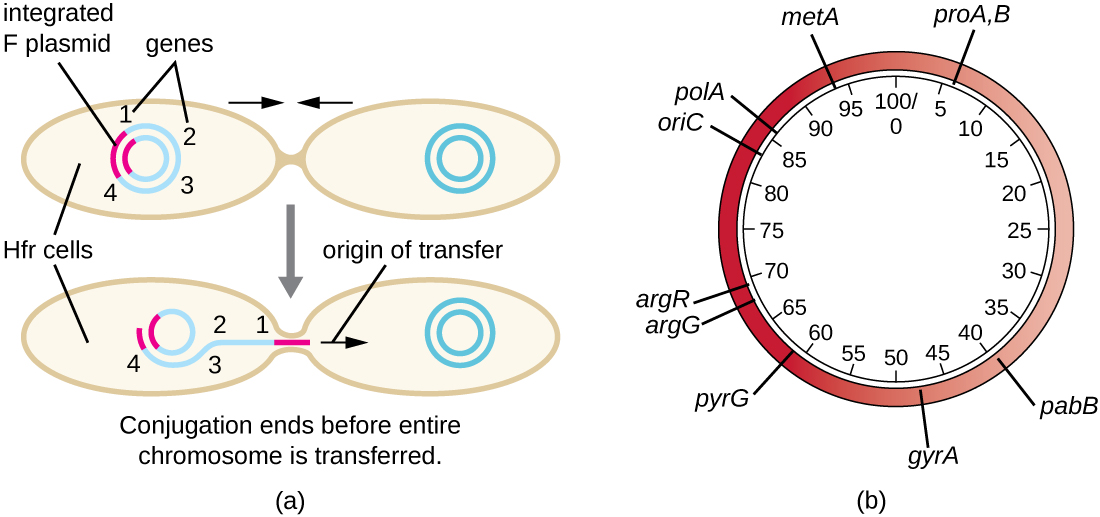
Consecuencias y aplicaciones de la conjugación
Los plásmidos son un tipo importante de elemento extracromosómico de ADN en bacterias y, en aquellas células que los albergan, se consideran parte del genoma bacteriano. Desde una perspectiva clínica, los plásmidos suelen codificar genes involucrados en la virulencia. Por ejemplo, los genes que codifican proteínas que hacen que una célula bacteriana sea resistente a un antibiótico particular se codifican en plásmidos R. Los plásmidos R, además de sus genes para resistencia antimicrobiana, contienen genes que controlan la conjugación y transferencia del plásmido. Los plásmidos R son capaces de transferirse entre células de la misma especie y entre células de diferentes especies. Los plásmidos R individuales contienen comúnmente múltiples genes que confieren resistencia a múltiples antibióticos.
Los genes requeridos para la producción de diversas toxinas y moléculas importantes para la colonización durante la infección también se pueden encontrar codificados en plásmidos. Por ejemplo, las cepas productoras de verotoxina de E. coli (VTEC) parecen haber adquirido los genes que codifican la toxina Shiga de su pariente gramnegativa Shigella dysenteriae a través de la adquisición de un plásmido grande que codifica esta toxina. El VTEC causa enfermedad diarreica grave que puede resultar en el síndrome urémico hemolítico (SUH), que puede conducir a insuficiencia renal y muerte.
En entornos no clínicos, los genes bacterianos que codifican enzimas metabólicas necesarias para degradar compuestos atípicos especializados como los hidrocarburos aromáticos policíclicos (HAP) también se codifican frecuentemente en plásmidos. Adicionalmente, ciertos plásmidos tienen la capacidad de pasar de células bacterianas a otros tipos de células, como las de plantas y animales, a través de mecanismos distintos de la conjugación. Dichos mecanismos y su uso en ingeniería genética están cubiertos en Aplicaciones Modernas de la Genética Microbiana.
Ejercicio\(\PageIndex{5}\)
- ¿Qué tipo de replicación ocurre durante la conjugación?
- ¿Qué ocurre para producir una célula de E. coli Hfr?
- ¿Qué tipos de rasgos se codifican en los plásmidos?
Transposición
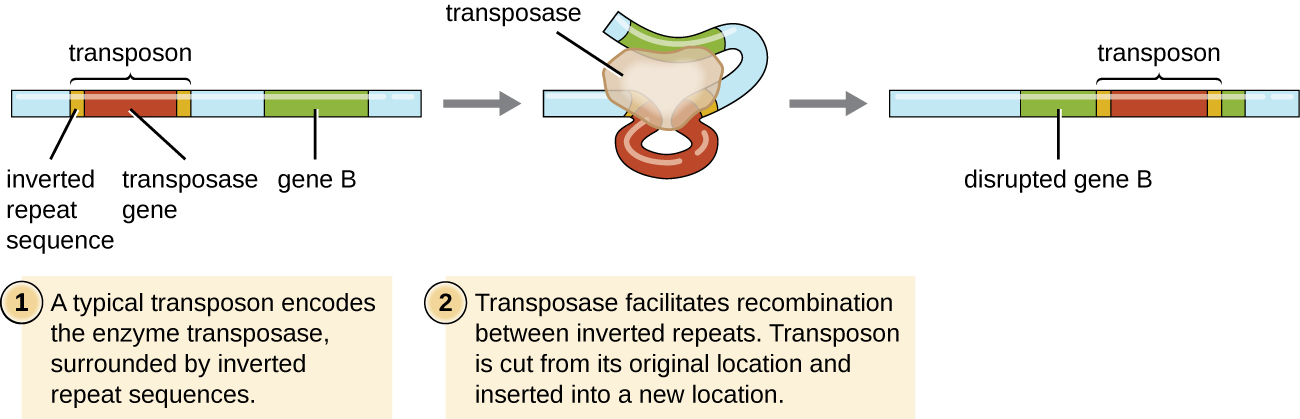
Los elementos genéticos llamados transposones (elementos transponibles), o “genes saltadores”, son moléculas de ADN que incluyen secuencias especiales de repetición invertida en sus extremos y un gen que codifica la enzima transposasa (Figura\(\PageIndex{6}\)). Los transposones permiten que toda la secuencia se extirpe independientemente de una ubicación en una molécula de ADN y se integre en el ADN en otra parte a través de un proceso llamado transposición. Los transposones fueron descubiertos originalmente en maíz (maíz) por la genetista estadounidense Barbara McClintock (1902—1992) en la década de 1940. Desde entonces se han encontrado transposones en todo tipo de organismos, tanto procariotas como eucariotas. Así, a diferencia de los tres mecanismos anteriores discutidos, la transposición no es procariota específica. La mayoría de los transposones son no replicativos, lo que significa que se mueven de una manera “cortar y pegar”. Algunos pueden ser replicativos, sin embargo, conservando su ubicación en el ADN mientras hacen una copia para insertarse en otro lugar (“copiar y pegar”). Debido a que los transposones pueden moverse dentro de una molécula de ADN, de una molécula de ADN a otra, o incluso de una célula a otra, tienen la capacidad de introducir diversidad genética. El movimiento dentro de una misma molécula de ADN puede alterar el fenotipo al inactivar o activar un gen.
Los transposones pueden llevar consigo genes adicionales, trasladándolos de un lugar a otro con ellos. Por ejemplo, los transposones bacterianos pueden reubicar genes de resistencia a antibióticos, moviéndolos de cromosomas a plásmidos. Se ha demostrado que este mecanismo es responsable de la colocalización de múltiples genes de resistencia a antibióticos en un solo plásmido R en cepas de Shigella causantes de disentería bacteriana. Dicho plásmido R se puede transferir fácilmente entre una población bacteriana a través del proceso de conjugación.
Ejercicio\(\PageIndex{6}\)
¿Cuáles son las dos formas en que un transposón puede afectar el fenotipo de una célula a la que se mueve?
| Término | Definición |
|---|---|
| Conjugación | Transferencia de ADN por contacto directo usando un pilus de conjugación |
| Transducción | Mecanismo de transferencia horizontal de genes en bacterias en las que se transfieren genes a través de infección viral |
| Transformación | Mecanismo de transferencia génica horizontal en el que el ADN ambiental desnudo es captado por una célula bacteriana |
| Transposición | Proceso mediante el cual el ADN se escinde independientemente de una ubicación en una molécula de ADN y se integra en |
Enfoque Clínico: Parte 3
A pesar del tratamiento antibiótico continuado, la infección de Mark continuó progresando rápidamente. La región infectada siguió expandiéndose, y hubo que ponerle un ventilador para que le ayudara a respirar. El médico de Mark ordenó la extirpación quirúrgica del tejido infectado. Después de una cirugía inicial, la herida de Mark fue monitoreada diariamente para asegurar que la infección no regresara, pero continuó propagándose.
Después de dos rondas adicionales de cirugía, la infección finalmente pareció estar contenida. Unos días después, Mark fue retirado del ventilador y pudo respirar por su cuenta. No obstante, había perdido una gran cantidad de piel y tejidos blandos en la parte inferior de la pierna.
Ejercicio\(\PageIndex{7}\)
- ¿Por qué la extirpación de tejido infectado detiene la infección?
- ¿Cuáles son algunas complicaciones probables de este método de tratamiento?
Conceptos clave y resumen
- La transferencia horizontal de genes es una forma importante para que organismos que se reproducen asexualmente como los procariotas adquieran nuevos rasgos.
- Hay tres mecanismos de transferencia horizontal de genes típicamente utilizados por las bacterias: transformación, transducción y conjugación.
- La transformación permite que las células competentes capten el ADN desnudo, liberado de otras células al morir, en su citoplasma, donde puede recombinarse con el genoma del huésped.
- En la transducción generalizada, cualquier fragmento de ADN cromosómico puede transferirse por empaquetamiento accidental del cromosoma huésped degradado a una cabeza de fago. En la transducción especializada, solo el ADN cromosómico adyacente al sitio de integración de un fago lisogénico puede transferirse como resultado de la escisión imprecisa del profago.
- La conjugación está mediada por el plásmido F, que codifica un pilus de conjugación que pone en contacto una célula F + que contiene plásmido F con una célula F.
- La rara integración del plásmido F en el cromosoma bacteriano, generando una célula Hfr, permite la transferencia de ADN cromosómico del donante al receptor. Adicionalmente, la escisión imprecisa del plásmido F del cromosoma puede generar un plásmido F' que puede transferirse a un receptor por conjugación.
- La transferencia de conjugación de plásmidos R es un mecanismo importante para la propagación de la resistencia a antibióticos en comunidades bacterianas.
- Los transposones son moléculas de ADN con repeticiones invertidas en sus extremos que también codifican la enzima transposasa, permitiendo su movimiento de una ubicación en el ADN a otra. Aunque se encuentran tanto en procariotas como eucariotas, los transposones son clínicamente relevantes en patógenos bacterianos para el movimiento de factores de virulencia, incluyendo genes de resistencia a antibióticos.