
10.3: Hidrólisis de fosfatos
- Page ID
- 2386

While kinase enzymes catalyze the phosphorylation of organic compounds, enzymes called phosphatases catalyze dephosphorylation reactions. Serine phosphatase, for example, catalyzes the following dephosphorylation of phosphoserine residues:
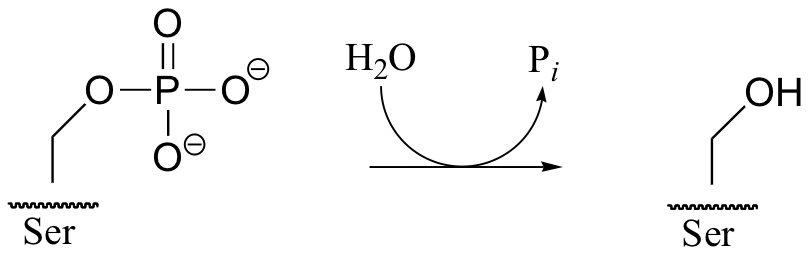
The reactions catalyzed by kinases and phosphatases are not the reverse of one another: kinases transfer phosphoryl groups from ATP (or sometimes other nucleoside triphosphates) to various organic compounds, while phosphatases transfer phosphoryl groups from organic compounds to water, which is a hydrolysis reaction.
Look again the serine phosphatase reaction. Two very different things could be happening, given the products that result. The reaction could be a phosphoryl transfer reaction, in which a phosphorus-oxygen bond is broken (reaction A). Alternatively, the water could be attacking the carbon of the serine side chain, breaking a carbon-oxygen bond and expelling the phosphate group in a nucleophilic substitution reaction (reaction B).
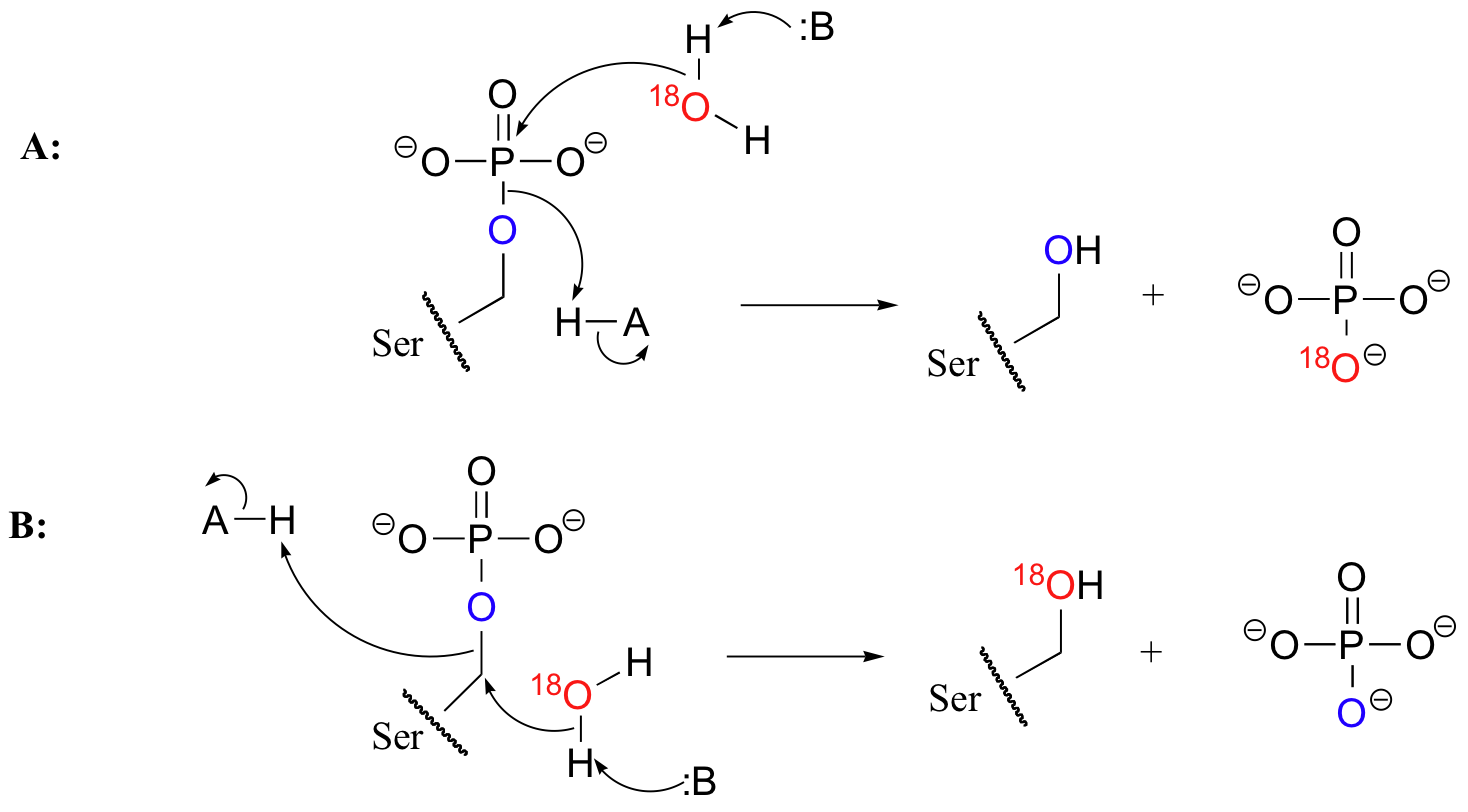
In order to find out which of these two mechanisms applies to phosphatases, scientists incubated a phosphatase enzyme with H218O, then used mass spectrometry to determine that the 18O ended up exclusively in the free phosphate (J. Biol. Chem. 1961 236, 2284). This result supports the idea that phosphatase reactions are phosphoryl transfer reactions (reaction A above), not nucleophilic substitutions.
Given this information, we would expect that the phosphatase reactions result in inversion of stereochemistry at the phosphate phosphorus. Experiments with chiral, 17O and 18O-labeled phosphate ester substrates show that this is indeed the case with many phosphatases (J. Am. Chem. Soc. 1993, 115, 2974). In other enzymes, however, dephosphorylation appears to occur with retention of configuration. Glucose-6-phosphate phosphatase is one such example (Biochem. J. 1982, 201, 665).
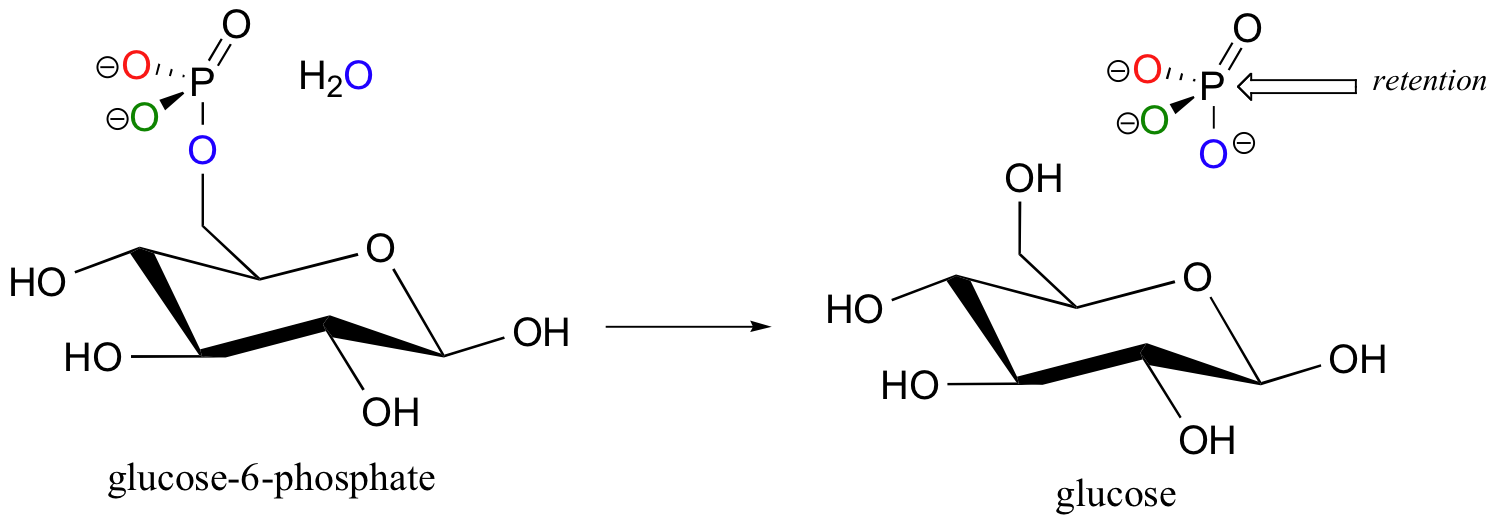
How could a phosphoryl transfer reaction result in retention of stereochemistry? Think back to the retaining glycosidase reaction (section 9.2), in which a nucleophilic substitution with overall retention of configuration was achieved via a double displacement mechanism. A nucleophile on the enzyme itself carries out the first nucleophilic attack (step 1) , then is subsequently displaced by water (step 2).
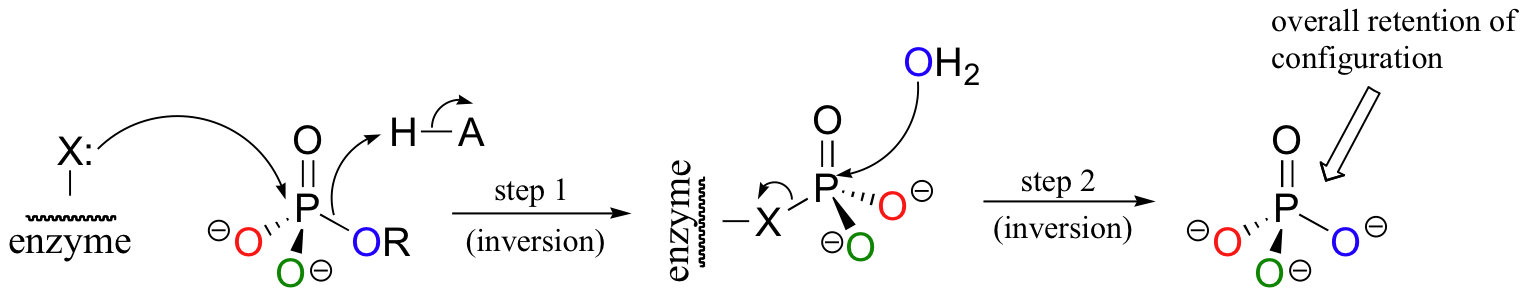
Could a similar thing be happening here? In order to find out, researchers ran the reaction with radioactive 32P-labeled glucose-6-phosphate, then stopped the reaction midstream in order to try to isolate the predicted enzyme-phosphate intermediate. The enzyme was chopped up into small pieces using proteases (we'll learn about these protein-chopping enzymes in chapter 12), and a radioactive phosphate was found to be covalently attached to an active site histidine (Biochem Biophys Acta 1972, 268, 698). Apparently, this histidine is acting in the role of the 'X' group in the figure above.