7.7: Los efectos del disolvente y el microambiente de los enzimas en la acidez
- Page ID
- 2366

Thus far in our discussion of acidity and basicity, we have not mentioned one very important factor– the effect of the solvent. The acidity of any compound, as we well know, depends on the difference in stability between the acid and its conjugate base. The ability of a carboxylic acid to donate a proton when in aqueous solution, for example, depends in large part on how the charged products that result from proton donation are stabilized through extensive hydrogen bonding interactions with the water solvent. The importance of the role played by solvent in this reaction becomes obvious when you consider that acetic acid, when reacting with water in the gas phase, has a measured pKa value greater than 100! (Recall that the pKa of acetic acid in water is 4.8).

Without the solvating, ion-stabilizing effect of solvent water, it is very difficult to pull apart the acetate anion and hydronium cation.
Because water is the solvent in biological chemistry, it is tempting to assume that solvent effects are a constant factor when considering the acidity or basicity of biomolecules. This is only partially correct. Keep in mind that virtually all biochemical reactions take place inside the active site pocket of an enzyme, rather than free in solution. The actual ‘microenvironment’ of an enzyme active site can often be very different from the environment outside in the ‘bulk solvent’. Consider, for example, the side chain carboxylate on an aspartate residue of an enzyme. The literature pKa of this carboxylic acid group is listed as 3.9, but this estimate assumes that the group is on the surface of the protein, exposed to solvent water. In the physiological buffer of pH ~ 7, a carboxylic acid group with pKa = 3.9 will be fully deprotonated and negatively charged. If, however, an aspartate side chain is buried deep inside the interior of the protein’s active site, and is surrounded primarily by hydrophobic residues such as alanine, phenylalanine, tryptophan, etc., the situation is very different.
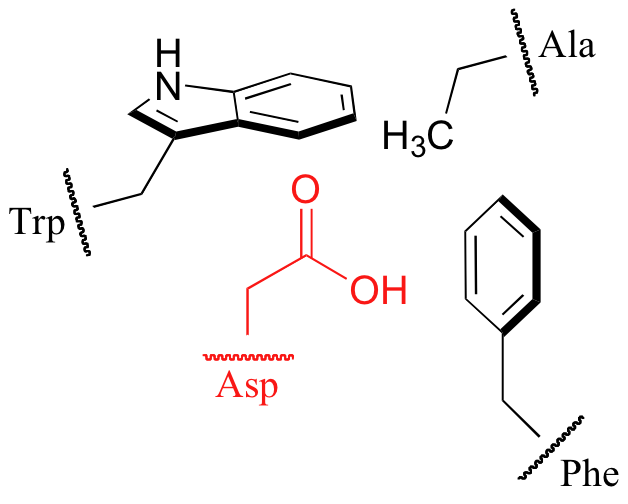
Cut off from the environment of the bulk solvent, the carboxylate group is now in a very hydrophobic microenvironment, a situation in which the protonated, uncharged state is stabilized relative to the deprotonated, negatively charged state. The overall effect is that the pKa for this aspartate residue is higher than 3.9 – it is less acidic, and more likely to be in its protonated form inside the protein.
A similar effect would be observed in a situation where the side chain carboxylate groups of two aspartate residues are located in close proximity to one another. Two negatively charged groups close to each other represents a very high energy, repulsive situation, and this can be relieved if one of the two side chains is protonated.
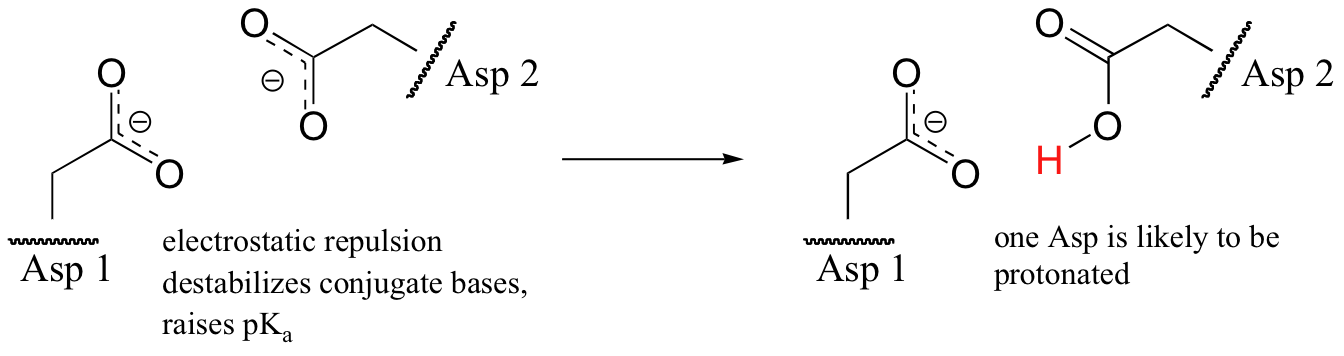
In this microenvironment, the proximity of one amino acid group directly effects the pKa of its neighbor.
Now consider a situation where a metal ion such as magnesium (Mg+2) or zinc (Zn+2) is bound in the interior of the enzyme, in close contact with an aspartate side chain. With a cation to interact with, the anionic, deprotonated state of the amino acid is stabilized, so the pKa of this Asp residue is likely to be substantially lower than 3.9.
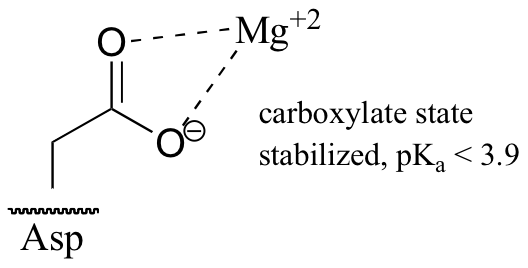
The metal ion in this situation is considered to be acting as a Lewis acid, (section 7.1B), accepting electron density from the carboxylate group.
The pKa-lowering effect of a metal cation can be dramatic – it has been estimated that a water molecule coordinated to a Cu+2 or Zn+2 ion can have a pKa as low as 7 (compare this to the ‘normal’ water pKa of 15.7!) (J. Am. Chem. Soc. 1984, 106, 630.)
Exercise 7.14: A lysine residue located deep in the interior of a protein is surrounded by alanines and leucines. In what direction will this alter the 'normal' pKa of the lysine side chain, and why?
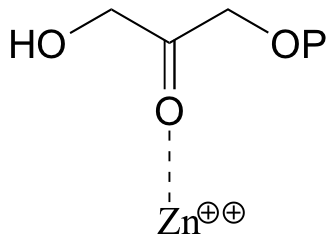
Exercise 7.15: In many biochemical reactions which involve the formation of an enolate intermediate, the carbonyl oxygen of the substrate is coordinated to a divalent metal ion (usually zinc or magnesium) in the active site. Explain, with structural drawings, how this ion-dipole interaction effects the acidity of the a-protons of DHAP.