
1.5: Estructura celular en la vida inanimada
- Page ID
- 56472

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)El descubrimiento de la primera célula suele atribuirse a Robert Hooke a mediados del siglo XVII. Pero la gente había observado células antes (huevos de pescado, huevos de ranas, huevos de aves). Lo significativo de la observación de Hooke fue que señaló que los organismos, al menos algunos de ellos, estaban compuestos por entidades más pequeñas a las que llamó células. H e pudo verlos en parte porque estaba usando el microscopio recientemente desarrollado. Pero lo que también hizo visibles las células fue que eran células vegetales, células que no sólo son más grandes que la mayoría de las células animales sino que también tienen paredes celulares que delinean cada célula y son rígidas y no colapsan cuando el material se corta en secciones delgadas. El material que estaba mirando era el corcho, la corteza exterior de un árbol, y lo que más obtenía eran los restos de células que ya no estaban vivas; su existencia se conservaba por sus gruesas y persistentes paredes celulares (Figura 1).

Si bien las observaciones de Hooke ilustraban que al menos algunas partes de algunos organismos estaban compuestas por unidades más pequeñas, el ion generalizat de que todos los organismos estaban compuestos de una a muchas 'unidades fundamentales', células, no iba a realizarse completamente por otros 200 años cuando se propuso y promovió la 'teoría celular'. Si bien la gente estaba muy consciente de los huevos de pescado y de rana, no habían apreciado que cuando los huevos se convirtieron en renacuajos el material se dividiera primero en muchas, muchas unidades pequeñas, cada una con la misma construcción básica. Con el tiempo, estas unidades se diversificaron entre sí y se transformaron en grupos para cumplir roles específicos. Estas están dispuestas de una manera particular para producir un todo complejo, un embrión de rana. Piensa en hacer castillos a partir de legos. Una transformación similar ocurre a medida que el óvulo fertilizado de las plantas se transforma en un embrión vegetal, aunque el óvulo de las plantas (aproximadamente 20 um) es 100 veces más pequeño que los de las ranas y aproximadamente cinco veces más pequeños que los de los humanos.
Uno de los grandes temas de la biología es la unidad y diversidad de la vida: cómo los organismos pueden ser simultáneamente tan similares y tan diferentes. El mismo concepto también se aplica a las células y es fundamental para la 'teoría celular': los seres vivos están formados por células, cuya organización básica, composición química y organización es similar; pero estas células también son diversas en términos de forma y función y la diversidad se ve tanto entre diferentes organismos (las células vegetales son diferentes de las células animales) pero también dentro de organismos (multicelulares).
La diversidad de células dentro de un organismo es uno, y para algunas personas el único, criterio para un organismo multicelular, no simplemente que estén hechas de múltiples células sino que al menos algunas de las células difieran entre sí. Antes de describir la diversidad de tipos celulares que se encuentran en algunos de los organismos que cubrimos, consideraremos criterios adicionales que podrían ser utilizados para definir la multicelularidad.
TEMAS
- Multicelularidad
- Estructura Celular de Hongos
- Quitridos
- Hongos coenocíticos filamentosos — moldes de pan y Glomeromycota
- Hongos septados — Hongos club y saco
- Estructura celular en moldes de agua
- Estructura celular en algas
- Algas pardas
- Algas Rojas
- Algas verdes
Multicelularidad

¿Un organismo unicelular puede ser multicelular? Uno podría pensar que esto es absurdo, sin embargo, puede ser cierto, dependiendo de las definiciones. Un caso ilustrativo es el Apicomplexa, grupo que incluye es el organismo causante de la malaria, Plasmodium. Los miembros de este grupo son siempre unicelulares pero tienen múltiples tipos celulares (Figura 2), células que se ven diferentes, se comportan de manera diferente y viven en diferentes lugares. La mayoría de los biólogos no considerarían el Apicomplexa como multicelular y preferirían considerarlo como una entidad biológica con múltiples formas, pero en un sentido del desarrollo plantea la misma pregunta que plantea la multicelularidad: si una célula es idéntica genéticamente a otra célula, ¿cómo pueden terminar buscando y comportarse de manera diferente?
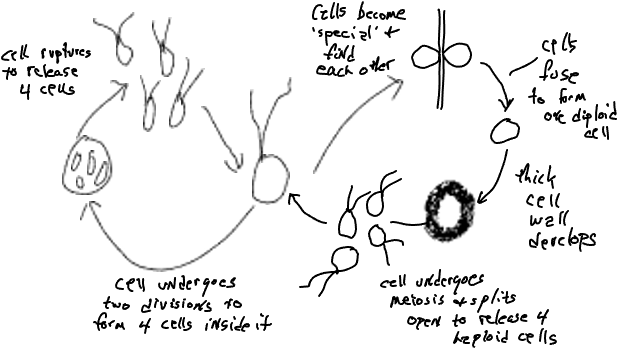
Si estudias cuidadosamente el ciclo de vida de Plasmodium notarás que parte de la diversidad celular está asociada con la reproducción y el proceso sexual, tema que se tratará con más detalle más adelante pero por ahora simplemente anota que la mayoría de los organismos producen diferentes células que están asociadas con la reproducción y/o el sexo. Las clamidomonas, un alga verde unicelular (Figura 3), produce células bi-flageladas que todas se ven y se comportan igual excepto por: (1) algunas células se ven diferentes porque pierden sus flagelas y se dividen dos veces para producir cuatro células mientras aún están dentro de la célula parental. Eventualmente se digiere la pared celular parental y se liberan las cuatro células de aspecto típico, (2) algunas células se ven normales pero son 'especiales', funcionalmente diferentes, porque son capaces de fusionarse con otras 'células especiales'. Las células se vuelven 'especiales' debido a las condiciones cambiantes, por ejemplo, los nutrientes se vuelven menos disponibles. (3) la célula formada por la fusión de dos células especiales se desarrolla en una célula de aspecto muy diferente, una sin flagelos y una pared celular gruesa. También funciona de manera diferente, siendo metabólicamente inactivo. Eventualmente se activa y se divide para producir cuatro células hijas de aspecto típico (bi-flageladas) que se liberan después de digerir y debilitar la pared celular. Los tres tipos de células atípicas están asociados con el sexo y el proceso de meiosis (lado derecho del diagrama anterior) se discutirá con más detalle más adelante, junto con una consideración de la reproducción (lado izquierdo del diagrama).
Obviamente muy pocos considerarían que organismos unicelulares como Plasmodium o Chlamydomonas son multicelulares a pesar de que pueden exhibir múltiples tipos celulares. El punto es que la posesión de células de diferente aspecto o de diferente comportamiento (por ejemplo, óvulo, esperma, cigoto, espora) que se asocian con el proceso reproductivo o sexual no son suficientes para etiquetar a un organismo como multicelular. Esto es trivialmente cierto para los organismos unicelulares pero también para los organismos coloniales que están formados por múltiples células que son casi todas iguales y es la razón de la distinción entre 'multicelularidad simple' y 'multicelularidad compleja'.

Consideremos las algas verdes filamentosas Ulothrix, (Figura 4) que existe como cadenas de células, la mayoría de las cuales todas se parecen (ver la parte central del dibujo anterior, etiquetada como 'a'), pero a veces algunas de las células se transforman (es decir, se desarrollan) para producir células móviles que tienen cuatro flagelos ('c' en el diagrama) y se liberan del filamento. Estas células pueden establecer nuevos filamentos uniéndose a un sustrato y dividiendo para producir nuevos filamentos ('d'). Otras células transforman y producen y posteriormente liberan células con solo dos flagelos ('f'). Estas células deben encontrar otra célula bi-flagelada, fusionarse con ella ('g'), luego someterse a meiosis para formar cuatro células cuadriflageladas ('h') que pueden unirse a un sustrato y formar nuevos filamentos.
Otro ejemplo de multicelularidad simple con diversos tipos celulares asociados a reproducción y sexo se encuentra en Oedogonium, (Figura 5) otra alga verde filamentosa. La mayoría de las celdas son cilíndricas con longitudes aproximadamente cinco veces más largas que su ancho. Pero también hay óvulos, que son más cortos y se abultan en el medio y células productoras de esperma mucho más cortas, con longitudes iguales o más cortas que su anchura. Ambos tipos de células desarrollan agujeros en sus paredes que permiten que las células cortas liberen espermatozoides móviles y permiten que los espermatozoides entren en los óvulos dando como resultado la producción de células con información genética de dos padres. Esto se convierte en una espora latente que finalmente se libera del filamento. Un tipo celular adicional puede desarrollarse a partir de las células normales a medida que desarrollan flagelos y eventualmente se liberan del filamento como células capaces de formar nuevos individuos.

Estos ejemplos demuestran tipos celulares comunes asociados al sexo y reproducción y encontrados en organismos con crecimiento colonial (mostrando 'multicelularidad simple'): (1) células que son capaces de fusionarse entre sí, llamadas gametos que a veces se diferencian en óvulo y esperma, (2) cigoto, una célula formada por la fusión de gametos, (3) zoosporas, células flageladas especializadas para la movilidad (dispersión en el espacio) (e.g., la célula marcada 'c' en el diagrama de Ulothrix), (4) células inactivas, inactivas, latentes que permiten la dispersión en el tiempo. En general, estas se llaman esporas (pero tenga en cuenta que las zoosporas NO están inactivas). Estas células inactivas se denominan endoesporas en bacterias y akinetes en cianobacterias. En una serie de organismos (por ejemplo, Chlamydomonas, Oedogonium, moho del pan) las esporas se desarrollan a partir de cigotos y se llaman cigoesporas. Veremos una serie de esporas diferentes en los hongos.

Para repetir, ninguno de los ejemplos considerados hasta ahora sería considerado multicelular a pesar de que producen múltiples células, algunas de las cuales son diferentes estructural y funcionalmente de otras. ¿Qué características adicionales contribuyen al caso de la 'multicelularidad' verdadera? Una característica es la dependencia mutua. La mayoría de las células de Anabena, una cianobacteria, tienen clorofila y fotosintetizan para alimentarse ellas mismas. Anabena también produce células llamadas heterocistas que carecen de clorofila y son incapaces de fotosintetizar. Son 'alimentados' por celdas adyacentes, 'regulares'. Los heterocistos son significativos porque pueden tomar gas dinitrógeno, una forma de nitrógeno que las células 'regulares' no pueden utilizar, y convertirlo en una forma que las células regulares pueden utilizar (más detalles en el capítulo 22). De ahí que los heterocistos y las células regulares dependan unos de otros. Al menos algunos trabajadores considerarían la dependencia de un tipo de célula sobre otro como un criterio que hace multicelular a Anabena. Otra característica que Anabena demuestra y que es significativa para el concepto de multicelularidad es el movimiento material (alimento, compuestos nitrogenados) entre las células del organismo. Esto realmente es una forma de comunicación intercelular, y muchos trabajadores consideran que la comunicación intercelular es un requisito clave que define la multicelularidad. Las características que potencian la comunicación incluyen conexiones transcelulares, plasmodesmas en el caso de las plantas.
Otro criterio que se ha aplicado para definir la 'verdadera multicelularidad' es una organización celular tridimensional, es decir, no solo celdas agregadas en una dimensión (un filamento) o agregadas en dos dimensiones (una lámina), o agregadas en tres dimensiones sin un programa de desarrollo determinado. La verdadera multicelularidad para algunos trabajadores requiere de un programa de desarrollo que produzca una forma tridimensional distinta que requiera que el tamaño, la forma y las actividades de las células estén influenciadas por una combinación de ubicación (dónde están en el organismo) y de dónde provienen (herencia celular). Tal estructura requiere una amplia comunicación y coordinación entre las células. Tenga en cuenta, aunque no necesariamente requiere de tejidos conformados por diferentes tipos de células.
Los trabajadores que estudian la evolución de la vida creen que la multicelularidad simple (colonias) ha aparecido al menos veinticinco veces. Multicelularidad compleja (o lo que algunos llamarían multicelularidad 'verdadera'), pero definida sin el requerimiento de múltiples tipos de células y tejidos, ha evolucionado al menos diez veces: una vez en el grupo que condujo a los animales, una vez en el grupo de algas verdes que condujo a las plantas, una vez en otro grupo de algas verdes que lo hizo no conducen a plantas, dos veces en las algas rojas, dos veces en las algas pardas, tres veces en los hongos, y varias veces en los procariotas. Si la definición de multicelularidad requiere tejidos con múltiples tipos celulares (no asociados con el sexo y la reproducción) entonces el número de grupos multicelulares se reduce mucho: solo animales, plantas y muy pocos hongos.
A continuación se describe la estructura celular de grupos selectos de organismos inanimados, organismos que podrían considerarse multicelulares. Se excluyen las plantas vasculares; estas se consideran en el siguiente capítulo.
Estructura Celular en Hongos
Si bien muchos biólogos caracterizan al grupo fúngico como 'multicelular', muchos claramente no lo son. Tres grupos fúngicos son coenocíticos y ninguno de ellos se caracteriza fácilmente por ser multicelulares. Y los hongos unicelulares, que existen en varios grupos fúngicos, ciertamente no serían considerados multicelulares. Los hongos que podrían considerarse poseedores de multicelularidad compleja lo hacen solo por una pequeña porción de su existencia y solo en una pequeña porción de su estructura. Casi todos los hongos producen una estructura filamentosa, una hifa (hifas plurales), que difícilmente es multicelular y se describe mejor como colonial, un organismo filamentoso sin especialización celular alguna. En tres de los grupos fúngicos las hifas carecen de paredes cruzadas y son coenocíticas. Las hifas crecen desde la punta y pueden ramificarse en dos. La ramificación también puede ocurrir ocasionalmente lejos de la punta. Las hifas también pueden fusionarse entre sí, produciendo así una estructura anastomosante. La red de hifas así formada se llama micelio. El micelio típico es una estructura de alimentación con una forma dinámica (creciendo y muriendo simultáneamente), difusa, muy adecuada para obtener nutrientes: posee tanto una gran superficie de absorción como también penetra en un gran volumen ambiental, lo que le permite 'extraer' un sustrato para nutrientes. El micelio de alimentación es el sistema digestivo de los hongos y simultáneamente sirve como: (1) una estructura para digerir los alimentos, (2) un intestino delgado para absorber nutrientes (3) un sistema circulatorio para mover los nutrientes a otras partes del hongo y finalmente a las estructuras reproductivas. Obsérvese que ninguna de estas funciones está asociada con tipos celulares, tejidos u órganos específicos.

Dos grupos fúngicos se consideran celulares porque tienen una especie de paredes cruzadas. Estos se llaman septos (septo singular) y solo sellan parcialmente las células porque están perforadas por poros que son lo suficientemente grandes como para permitir que muchos materiales (ribosomas, mitocondrias y a veces núcleos) se muevan de una célula a otra. Así, la composición de estos hongos se encuentra entre coenocítica y celular. A continuación se describe la estructura celular de grupos específicos de hongos.
Quitridos
El grupo fúngico considerado más primitivo (probablemente mejor descrito como el grupo que divergió en el momento más temprano del resto de hongos) son los Chytridiomycota (quítridos). Los quítridos son pequeños organismos acuáticos, tanto marinos como de agua dulce, con algunos que existen en la película de agua que rodea las partículas del suelo. La mayoría son microscópicas, generalmente del tamaño de un organismo unicelular típico (menos de 100 um). Pueden ser verdaderamente unicelulares (un núcleo) o coenocíticos (núcleos múltiples). Muchos producen formas que son aproximadamente esféricas. Varios quítridos forman lo que se llaman rizoides, extensiones similares a raíz del cuerpo principal que los unen a una fuente de nutrición, a menudo polen, las esporas de otros organismos, o una sola célula de un organismo multicelular vivo o muerto. Además de la unión, los rizoides también sirven para aumentar la superficie y permitir una digestión y absorción más efectivas. Algunos quítridos forman filamentos (hifas) que ocasionalmente se ramifican para formar un micelio muy pequeño. El grupo se distingue de todos los demás hongos por tener esporas flageladas, móviles.
Hongos filamentosos coenocíticos
Dos grupos fúngicos adicionales, los mohos del pan (Zygomycota) y el Glomeromycota, son sustancialmente más grandes que los quítridos y producen hifas coenocíticas y forman micelios. Si bien ambos grupos carecen de tipos celulares específicos, algunos sí forman estructuras específicas que se podrían denominar órganos, excepto que son partes de células, no compuestas por células. Para los moldes de pan estas estructuras, estolones, rizoides y esporangióforos, son relativamente grandes y se pueden ver a simple vista. En la Glomeromycota los 'órganos' son estructuras mucho más pequeñas, pero nuevamente son partes de células, no células. Los Glomeromycota son los hongos que, junto con las plantas, forman endomicorrizas, asociaciones entre raíces de plantas y hongos. En las endomicorrizas, el hongo penetra en las células radiculares individuales ('endo' significa en el interior) y dentro de estas células el hongo forma dos estructuras, arbusculos arbóreos y vesículas esféricas (los hongos endomicorrícicos a veces se llaman micorrizas vesiculares-arbusculares o simplemente micorrizas VA). Estas estructuras son extensiones fuera de las hifas coenocíticas. Los arbusculos son estructuras con algunas similitudes con los rizoides de los quítridos. No cumplen ningún papel de anclaje pero son significativos en el aumento del área de contacto entre el hongo y la planta, permitiendo así un mayor movimiento de material entre ellos. Las vesículas son cuerpos esféricos que se cree que almacenan materiales para el hongo y que pueden convertirse en esporas.
Hongos Septados - Hongos Club y Saco
Los dos grupos más grandes de hongos, hongos club (Basidiomycota) y hongos saco (Ascomycota) no son coenocíticos. Producen hifas que son septadas, con paredes transversales para delinear células individuales. A medida que se extiende la punta creciente de una hifa, las divisiones nucleares (mitosis) se coordinan con las divisiones celulares (citocinesis) que producen nuevas paredes transversales con un poro sustancial. Estas paredes transversales están orientadas perpendicularmente al eje largo de la hifa. Como resultado, nuevas células, cada una con un núcleo, se producen secuencialmente en la punta a medida que crece. La celda de punta también es capaz de dividirse ocasionalmente de manera que la punta se bifurque, se divida en dos, formando una rama. La ramificación a veces también ocurre lejos de la punta, resultado de una excrecencia de una célula previamente formada.
Bajo ciertas condiciones las hifas de Ascomycota y Basidiomycota también pueden formar micelios mucho más compactos, generalmente asociados con la reproducción, y algunos de los cuales se acercan en forma a estructuras que son más típicas de los organismos multicelulares. Estas estructuras pueden variar en organización desde una masa amorfa de hifas compactas (también conocido como micelio denso), denominado estroma, en el que se forman 'cuerpos fructíferos' (estructuras productoras de esporas) (ver enfermedad de la mancha alquitranada), hasta estructuras mucho más definidas y determinadas como un hongo típico con tallo y tapa. Por lo general, las esporas se producen a partir de una estructura que de alguna manera está elevada, lo que permite una mayor dispersión de esporas por el viento. La mayoría de estas estructuras productoras de esporas se llaman hongos si son visibles a simple vista; y la mayoría son hongos club (Basidiomycota) pero algunos son hongos de copa (Ascomycota). Dentro de los dos grupos hay una amplia variedad de formas corporales fructíferas, todas desarrollándose como resultado del patrón de crecimiento de múltiples hifas. Aunque estas estructuras están claramente organizadas y producen una forma final tridimensional consistente (determinada), generalmente muestran poca evidencia de tejidos o tipos celulares especializados, excepto aquellos directamente asociados a la reproducción sexual.
Otra estructura hifal densa y compacta producida por hongos septados se llama cordón fúngico o rizomorfo. Consiste en múltiples hifas paralelas y pegadas entre sí, formando un hilo que puede tener varios milímetros de diámetro (cf. a una hifa fúngica que típicamente es diez veces menor en diámetro que un cabello humano). Los rizomorfos son el resultado de una ramificación hifal repetida en ángulos muy bajos con respecto a la orientación de las hifas parentales, lo que permite que la rama permanezca estrechamente asociada y fusionada con la hifa que la produjo. El resultado es una estructura con poca capacidad de alimentación (o pérdida de agua) en comparación con un micelio difuso debido a su reducida superficie y menor penetración de su entorno. Pero el rizomorfo está especializado en la movilidad y permite que el hongo atraviese el espacio de bajos nutrientes o el espacio ambientalmente hostil (por ejemplo, seco) y potencialmente llegue a una ubicación más favorable. Cuando y si el rizomorfo llega a una zona rica en nutrientes, la forma de crecimiento vuelve a ser un micelio difuso, nuevamente adecuado para la absorción de nutrientes. La extensión de rizomorfos requiere el transporte de nutrientes para alimentar las puntas de las hifas (donde se produce el crecimiento) utilizando nutrientes que se están adquiriendo a cierta distancia de la punta. Esta función de transporte es atendida por hifas especializadas de gran diámetro, análogas a los tubos que se encuentran en algas pardas y plantas vasculares. Otro tipo celular especializado que se encuentra en los rizomorfos, y que se encuentra hacia el exterior de ellos, son las 'hifas de revestimiento' de paredes gruesas que tienen una estructura nuevamente similar a algunas células (fibras) que se encuentran en las plantas vasculares. Estos le dan al rizomorfo integridad estructural y hacen que sea menos probable que se corte, algo que es de mucha más consecuencia para un rizomorfo que para una hifa individual.
Algunos hongos producen estructuras llamadas esclerocios, nuevamente una densa masa de hifas, pero los esclerocios a menudo se desarrollan en formas distintas y a veces muestran especialización entre una 'corteza' externa dura y células internas. Las células en el interior a menudo tienen reservas sustanciales de alimentos y los esclerocios suelen ser estructuras que hibernan, estructuras que pueden volverse inactivas. El alimento almacenado permite que el esclerocio, que a veces se dispersa, sobreviva al invierno y reanude el crecimiento cuando regresan las condiciones favorables.
Estructura celular de moldes de agua
Los mohos de agua, o Oomycota, son similares en forma a los moldes de pan (Zygomycota), estando ambos grupos compuestos por filamentos coenocíticos, hifas, que colectivamente forman un micelio pero no muestran especialización celular más que las células asociadas a la reproducción. El grupo incluye algunos patógenos importantes de las plantas, como el tizón tardío de la papa, el síndrome de muerte súbita del roble y las enfermedades de 'amortiguación'. A pesar del nombre, no son mohos (hongos) y muchos son terrestres no acuáticos (aunque el representante probablemente más visto, al menos si tienes acuario, es una pelusa blanca que ocurre en peces muertos). Los mohos de agua alguna vez se agruparon con los hongos debido a su morfología, pero ahora se considera que están en el grupo heteroconte que también incluye algas pardas y diatomeas.
Estructura celular de moldes de limo celular
Durante la parte unicelular de la existencia del moho de limo celular obviamente no hay diferenciación celular, pero cuando ocurre la agregación las células adquieren diferentes destinos y uno podría considerar que esto refleja una multicelularidad compleja. En particular, y de significación evolutiva, solo algunas células del agregado terminan finalmente produciendo células reproductivas. Otras células, por ejemplo, las células en el tallo de la estructura productora de esporas, contribuyen al éxito reproductivo elevando la estructura productora de esporas pero no obtienen ventaja evolutiva por 'hacer un buen trabajo' ya que no están necesariamente representadas en la siguiente generación. Esto contrasta con la producción más normal de multicelularidad donde todas las células de un organismo multicelular son derivadas (y por lo tanto comparten una composición genética) con la 'célula iniciadora' original, típicamente un cigoto o una espora. Debido a su trayectoria relativamente inusual hacia la multicelularidad, los determinadores del destino celular en los mohos de limo celular son un área de estudio activo. No hay diferencias visuales/estructurales obvias entre las células de la babosa o la estructura fructífera pero sus posiciones son diferentes, algunas están en el exterior, otras dentro; algunas están cerca del frente, otras están detrás. Y la posición parece ser importante para determinar el destino celular, incluyendo la posición en el momento de la agregación. Además, existe una fuerte evidencia de que el destino celular y el 'conocimiento' de la posición, es el resultado de la comunicación a través de señales químicas, uno de los criterios que los trabajadores consideran como un aspecto definitorio de la multicelularidad compleja.
Estructura Celular de Algas
Las algas son una agrupación artificial de organismos eucariotas fotosintéticos acuáticos cuya clasificación permanece en algún flujo. Muchas algas son unicelulares o coloniales ('multicelularidad simple') y no muestran ninguna especialización celular excepto las células asociadas con el sexo y la reproducción. La mayoría de estos organismos son pequeños, microscópicos o apenas visibles a simple vista. Pero hay tres grupos de algas, llamadas coloridas algas rojas, algas pardas y algas verdes, que incluyen formas que se describen como 'macroalgas'. Son grandes (es decir, no microscópicas, generalmente de al menos 0.1 m de extensión) y con una estructura organizada, a menudo compuesta por órganos: tallos y ramas cilindricos, cuchillas aplanadas, sacos de aire, retenedores en forma de raíz y otras características. Casi todas las algas pardas y la mayoría de las algas rojas podrían describirse de esta manera. Esta forma es proporcionalmente menos común en las algas verdes donde muchos miembros son más pequeños y donde un número considerable de formas son unicelulares. Usando las definiciones más indulgentes de multicelularidad, la mayoría de las macroalgas serían consideradas multicelulares. El uso de definiciones más restrictivas, en particular requiriendo conexiones intercelulares y comunicación que permita la cooperación entre células, reduce sustancialmente el número de macroalgas que se consideran “verdaderamente multicelulares”. Sin embargo, parece que la multicelularidad compleja rígidamente definida ha evolucionado independientemente al menos dos veces en cada uno de los tres grupos de macroalgas (junto con tres veces en los hongos, una vez en animales y dos veces en bacterias).
Algas pardas
Este grupo solía considerarse un filo pero ahora es considerado como una entidad más abajo en la escala taxonómica (una clase, o una familia, o un nivel intermedio). Si bien casi todas las algas pardas son macroalgas, muchas carecen de diferenciación celular y una organización tisular obvia. La estructura es básicamente filamentosa, con los filamentos ramificándose e interactuando entre sí para formar una estructura distinta, similar a la forma en que ciertos hongos producen hongos a partir de hifas. Parece que la multicelularidad 'verdadera', con presencia de diferenciación celular y tejidos, ha evolucionado dos veces en este grupo, una vez en una línea que incluye rockweed (Fucus) y otra en una línea que incluye algas gigantes (Laminaria). En ambos grupos hay distintos órganos (holdfast, stipe, blade, vejigas de aire) junto con cierta diferenciación celular y tisular; en particular, hay células especializadas para permitir el movimiento a larga distancia de carbohidratos por todo el organismo. Estas células son análogas (¡no homólogas!) con células transportadoras de plantas vasculares y tienen características similares: son células alargadas con diámetros grandes y con múltiples conexiones transcelulares entre células adyacentes. Otro tipo celular distinto, también análogo a lo que se encuentra en las plantas vasculares, son las células con paredes celulares engrosadas que proporcionan integridad estructural, permitiendo que un organismo grande se mantenga unido a pesar de ser empujado y arrastrado por las corrientes oceánicas y/o las olas.
Algas Rojas
El grupo de algas rojas tiene aproximadamente cuatro veces el número de especies que las algas pardas y es más diverso en términos de morfología. Un número sustancial son macroalgas con una estructura organizada definida. Como fue el caso de los hongos y algunas algas pardas, su complejidad tridimensional es consecuencia del crecimiento e interacciones de múltiples filamentos. Los tejidos se pueden describir en algunas formas, con las porciones cilíndricas ('tallos') teniendo distintas capas externas e internas y poseyendo células que se distinguen por su tamaño y pigmentación. Algunas algas rojas forman costras ('crustosas') y existen como recubrimiento en la superficie de diversos sustratos. En muchas formas crustosas existe una organización estratificada, con los filamentos próximos al sustrato teniendo celdas diferentes a las de arriba. Algunas de las algas rojas crustosas desarrollan una capa de células divisorias cerca de la superficie superior cuya acción permite que la corteza se espese y también permite el desarrollo de estructuras reproductivas.
Algas Verdes
De los tres grupos de algas que incluyen macroalgas, el grupo de algas verdes es, con mucho, el más grande (15, 000 especies), con muchas formas unicelulares, filamentosas simples y coloniales. En consecuencia, las macroalgas son una porción mucho menor del total de especies. Algunas de las macroalgas son sifonáceas, a menudo compuestas por filamentos multinucleados que interactúan para formar una forma tridimensional. La mayoría de las filogenias modernas (cladísticas) dividen las algas verdes en dos grupos, uno de los cuales, además de incluir (algunas) algas verdes, contiene todos los organismos considerados plantas. Ambos grupos exhiben una variedad de formas, incluyendo algunas macroalgas con multicelularidad 'compleja', mostrando crecimiento tridimensional, conexiones celulares y distintos tipos de células.