
1.9: Crecimiento secundario
- Page ID
- 56498

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)
El crecimiento pr imario extiende el eje raíz/brote y produce raíces de rama y brotes. Recordemos que el ancho de una raíz o brote producido por el crecimiento primario es limitado debido a que la mayoría de las células no se expanden mucho en la dirección radial. Y para la mayoría de las plantas la división celular en el meristema apical es casi exclusivamente en una dirección que provoca más células en el eje largo, con muy pocas divisiones que aumentarían el número de células a través del diámetro de la raíz o brote. El crecimiento primario permite que la planta se alargue y se vuelva 'más ocupada' (debido a las ramas agregadas) pero en general no permite que las raíces y los brotes se vuelvan muy anchos. Esta falta de crecimiento radial limita la altura de la planta, sin tallos más gruesos para resistir los esfuerzos combinados del viento y la gravedad, es difícil que una planta se vuelva alta. Al mismo tiempo, la competencia por la luz da una clara ventaja a las plantas más altas. Otro problema con solo tener crecimiento primario es que la fuente de agua (las partes más jóvenes de las raíces) sigue alejándose del lugar que necesita agua, las puntas de los brotes donde están las hojas y donde se produce un nuevo crecimiento. Si bien se pueden crear raíces de rama, o raíces adventicias para acortar la ruta, el hecho aún permanece que el crecimiento primario separa las fuentes de agua de las partes que necesitan agua. Adicionalmente, tanto las células conductoras del xilema como las del floema pueden fallar por diversas razones. Debido a que la reparación de las células existentes a menudo no es posible y debido a que el crecimiento primario no permite la producción de células conductoras de reemplazo, la capacidad de hacer que los tallos sean más anchos, y en particular hacerlos más anchos con la adición de celdas de transporte y células de soporte estructural, proporciona algunos claras ventajas, incluyendo pero no limitado a la longevidad.
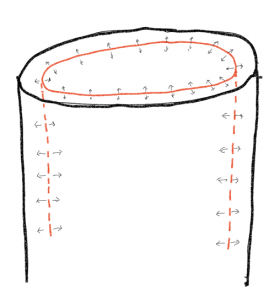
El crecimiento radial es posible en plantas que producen lo que se conoce como meristemos laterales. Estos meristemos son capaces de aumentar la circunferencia de raíces y brotes más allá de lo que se produce por el crecimiento primario. Los meristemos laterales son cilindros de células embrionarias que recorren toda la longitud del eje raíz/brote. La división celular en estas regiones embrionarias, seguida de la expansión de las nuevas células, permite que los tallos y las raíces aumenten de grosor en un tipo de crecimiento definido como crecimiento secundario. Debido a que cualquier expansión radial romperá los tejidos fuera de donde ocurre el crecimiento, el tejido dérmico producido en el crecimiento primario se va a abrir y se necesita producir una nueva 'piel'. En consecuencia, el crecimiento radial en raíces y tallos requiere de dos meristemas laterales, uno, el cámbium vascular, responsable de la mayor parte del aumento de la cincha, y uno, el cambium de corcho, responsable de hacer una nueva piel. A diferencia de las nuevas células producidas por los meristemos apicales, las divisiones celulares de los meristemos laterales son generalmente paralelas a la superficie de la raíz o brote y las nuevas células se expanden en dirección radial (interior/exterior), aumentando así el diámetro del tallo o raíz pero sin cambiar su longitud.
TEMAS
- Cambium vascular
- Xilema secundario y floema secundario
- Rayos
- Cambium de corcho
- Orígenes evolutivos del crecimiento secundario
- Monocotiledóneas anchas y madera
Cambium vascular
El cambium vascular produce nuevo tejido vascular y es responsable de la mayor expansión radial del mismo. En una sección transversal de un tallo o raíz el cámbium vascular existe como un círculo de células, solo unas pocas células de ancho. En tres dimensiones el cambium vascular es un cilindro. En el desarrollo, el cámbium vascular se origina a partir de células indiferenciadas localizadas entre el xilema y el floema que fueron producidas por el meristema apical. Recordemos que el crecimiento primario de tallos produce xilema y floema en haces que, para todos los grupos distintos de las monocotiledóneas (que no presentan crecimiento secundario), ocurren en un anillo dentro del tallo. Para hacer del cámbium vascular un anillo continuo requiere que las células entre los haces vasculares sean estimuladas para comenzar a dividirse. El cámbium vascular también puede desarrollarse en raíces, originándose nuevamente a partir de células localizadas entre el xilema y floema y células adicionales para formar un anillo continuo.
Xilema secundario y floema secundario
Las divisiones celulares del cámbium vascular producen xilema y floema que se denomina 'secundario' para distinguirlo del xilema primario y floema producido por los meristemos apicales. Si alguna célula en particular producida por la acción del cámbium vascular se diferencia en floema secundario o xilema secundario depende de su posición, un factor común que controla la diferenciación celular. En el caso más simple, cuando una célula del cambium vascular se divide produce una célula que permanece embrionaria (no se expande ni se diferencia) y una célula que está destinada a expandirse radialmente y diferenciarse. Si la célula en maduración está al exterior de la célula que permanece meristemática está destinada a convertirse en una célula de floema: un miembro de tubo de tamiz, una célula de parénquima o una fibra. Si la célula en maduración se produce al interior de la célula que permanece meristemática está destinada a convertirse en una célula de xilema: un elemento tubular vascular o una traqueidea o una fibra o una célula de parénquima. La mayoría de las nuevas células producidas por el cambium vascular están en el interior, por lo que se produce más xilema secundario que floema secundario.

La gran mayoría de las células producidas por el cámbium vascular son alargadas a lo largo del eje largo del tallo (fibras, elementos de tubo de tamiz, células de tamiz en el floema; fibras, traqueides, miembros de tubo de vaso en el xilema). Esta forma no es el resultado del crecimiento de estas células; cualquier elongación de estas células en la dirección arriba/abajo es imposible: un tallo leñoso no puede alargarse en el medio, solo desde la punta. La forma alargada de estas células es el resultado de la forma de la célula que se dividió para producirlas. El cámbium vascular consiste principalmente en células, llamadas iniciales fusiformes, que son alargadas y que, después de dividirse, producen células hijas que también son alargadas. Estas células sólo se expanden en dirección radial, es decir, engordan, no más, produciendo un tallo que es más ancho, no más alto.

Rayos
Sin embargo, hay algunas células del cámbium vascular, llamadas iniciales de rayos, que no son alargadas sino que son más o menos cúbicas y producen células de parénquima que no son alargadas en dirección arriba/abajo sino que son ligeramente alargadas en dirección radial. Las células rectangulares del parénquima producidas por las iniciales de los rayos se encuentran en racimos (es decir, es probable que una inicial de rayo tenga una inicial de rayo por encima y/o por debajo de ella en el cámbium vascular), y forman estructuras llamadas rayos que discurren radialmente desde el interior hacia el exterior del tallo. Los rayos van desde una célula en grosor y menos de 10 celdas de altura (es decir, a lo largo del eje longitudinal de la raíz/tallo) e invisibles a simple vista, hasta rayos que son cientos de células en altura y decenas de células en grosor y fácilmente visibles a simple vista. Los rayos se producen tanto en el xilema secundario como en el floema secundario y son particularmente significativos para el almacenamiento de carbohidratos. Los carbohidratos transportados por el floema se almacenan en rayos y luego se pueden movilizar cuando sea necesario. En el xilema secundario los rayos también son significativos ya que son las únicas células vivas presentes debido a que las otras células secundarias del xilema (fibras, traquieds y elementos de tubo vascular) mueren muy poco después de ser producidas. Si bien las células de los rayos no viven para siempre, sí viven por varios años, y además del almacenamiento de carbohidratos pueden responder a patógenos. Finalmente, cuando mueren, producen compuestos antimicrobianos/antifúngicos que permean los tejidos circundantes, generalmente oscureciéndolos y produciendo lo que se describe como duramen en la parte central de un tallo leñoso. El cilindro de xilema secundario aún con células vivas del parénquima se denomina albura y generalmente es de color más claro.

Madera

En casi todas las plantas, las células del xilema que son producidas por el cámbium vascular, denominadas xilema secundario, tienen una pared celular secundaria sustancial que contiene lignina y están fuertemente unidas a las células adyacentes. Así, este crecimiento secundario es un tejido estructuralmente fuerte y rígido y lo conocemos como madera, un material que es de mucha utilidad por sus características m ecánicas y también por su belleza.
En muchas regiones del globo la acción del cambium vascular es estacional, por ejemplo, solo ocurre en la primavera y principios del verano. A menudo, la naturaleza de las células del xilema producidas por el cámbium vascular varía estacionalmente de manera característica. Por ejemplo, es bastante común que las células producidas a finales de la temporada tengan lúmenes más pequeños y paredes celulares proporcionalmente más gruesas que las células producidas a principios de la primavera. Otro patrón común es el resultado de que los elementos de tubo de vaso solo se producen a principios de la primavera, de manera que el crecimiento de cada primavera se identifica fácilmente por la presencia de elementos tubulares grandes. En consecuencia, generalmente existe un contraste sustancial entre las últimas celdas formadas al final del verano y las primeras celdas formadas la primavera siguiente. Esto da como resultado un patrón conocido como anillos anuales cuando la madera se ve en sección transversal. En una sección longitudinal los anillos de crecimientoestán presentes como líneas paralelas.




Peridermo
A medida que las nuevas células producidas a partir del cambium vascular se expanden, la fuerza del xilema secundario es suficiente para evitar que se aplaste. En cambio, la expansión de nuevas células de xilema empuja hacia afuera y aplasta la mayoría de las células hacia el exterior del cámbium vascular. Las células del floema secundario recién producidas, siempre que estén vivas, pueden resistir ser aplastadas, al igual que las células de fibra altamente lignificadas que a menudo están presentes, pero la mayoría de las otras células son aplastadas por la expansión externa causada por el crecimiento de las células producidas por el cámbium vascular. Adicionalmente, este crecimiento externo, rompe la epidermis, la 'piel' original del tallo que fue producida por el meristema apical. Las plantas con crecimiento secundario producen un meristemo lateral adicional, el corcho-cambium, que produce células que forman una nueva piel llamada peridermo.
A diferencia del cambium vascular, el cambium de corcho generalmente no es un cilindro continuo. En cambio, generalmente existe como una serie de arcos que colectivamente forman un anillo. Al igual que el cambium vascular, el cambium de corcho produce diferentes células al interior y al exterior; las células producidas al exterior son de corta duración y tienen paredes celulares que están impregnadas con suberina, un compuesto impermeable. Las células que se producen al interior son células del parénquima y viven más tiempo, generalmente varios años. Esto es significativo porque las células del cámbium del corcho, a diferencia de las del cámbium vascular y los meristemos apicales, no son de larga vida; mueren a los pocos años, y se forma un nuevo cambium de corcho al interior, originado en las células del parénquima que se formaron al interior del corcho más viejo cambium. En los brotes, el cambium inicial del corcho se origina dentro de la corteza y los posteriores cambios de corcho se originan a partir de derivados de los anteriores. En las raíces el cámbium del corcho se origina en la actividad del periciclo y nuevamente se reforma hacia adentro a partir de las células del parénquima producidas por los cambios de corcho anteriores. Así, a través del tiempo el cámbium del corcho se mueve hacia adentro mientras todo el tallo crece hacia afuera debido a la acción del cambium vascular. Las células que produce el cámbium del corcho, así como las células del floema secundario, se comprimen continuamente por la expansión desde dentro; estos tejidos también se están separando a medida que aumenta la contorno del tronco.
En general, las células producidas al exterior por el cámbium de corcho están estrechamente empaquetadas y no tienen grietas ni espacios de aire, como fue el caso de la epidermis original. Pero el cambium de corcho produce regularmente áreas llamadas lenticelas donde hay grietas y fisuras. Se piensa que las lenticelas son significativas para permitir la penetración de oxígeno en el tallo. Al mismo tiempo, las lenticelas proporcionan un espacio para permitir el escape del agua y la entrada de patógenos. Esto vuelve a resaltar el hecho de que si bien algunos aspectos de la vida se ven favorecidos por el aislamiento del entorno exterior, otros aspectos de la vida requieren conexión con el ambiente exterior.

En los tallos leñosos el material al interior del cámbium vascular (todo ello xilema secundario, ignorando la pequeña porción de xilema primario y médula que puede permanecer en el centro del tallo) se llama madera. Todo el material fuera del cambio vascular: floema secundario, el cámbium de corcho y los productos de la actividad del cambio de corcho de uno a muchos, además de pequeños trozos de floema primario, corteza y epidermis, se denominan colectivamente corteza. El aspecto de la corteza varía enormemente debido a las diferencias en el comportamiento del cambium de corcho.


Orígenes evolutivos del crecimiento secundario
El crecimiento secundario y junto con él, las plantas leñosas, arbóreas aparentemente se han originado en múltiples ocasiones: una vez en un grupo que contiene musgos clubes actuales, una vez en un grupo que contiene colas de caballo actuales; al menos una vez y probablemente varias veces en grupos extintos de plantas ('helechos semillas') que no son agrupadas con cualquiera de las plantas existentes con semillas, y una vez en el grupo que produjo todas las plantas semilleras existentes (plantas con flores, coníferas, cícadas, ginkgo y gnetófitos). Aunque el crecimiento secundario apareció varias veces, también ha desaparecido varias veces: ningún musgo o cola de caballo existente muestra crecimiento secundario y muchas plantas semilleras, en particular muchas plantas con flores, no muestran crecimiento secundario. Las formas leñosas extintas fueron muy significativas en el pasado, en particular en el Paleozoico tardío (hace 350-250 millones de años) cuando formaron extensos bosques cuya productividad se utiliza aún a través de yacimientos de carbón y gas.
Monocotiledóneas anchas y amader
Si la madera se define como xilema secundario y las monocotiledóneas no tienen crecimiento secundario entonces las monocotiledóneas no tienen madera, pero esto no impide que algunas monocotiledóneas sean leñosas, es decir, poseer tejidos lignificados. El crecimiento primario a menudo produce células lignificadas, pero generalmente no tejidos extensos que están lignificados. En la mayor parte del crecimiento primario hay relativamente pocas células leñosas, pero en algunas monocotiledóneas (por ejemplo, el bambú, que es una hierba) los tejidos primarios pueden ser bastante leñosos. Además, debido a que las monocotiledóneas carecen de crecimiento secundario y debido a que el crecimiento primario es generalmente limitado en dirección radial, las monocotiledóneas son Sin embargo, algunas monocotiledóneas muestran expansión radial prolongada en el crecimiento primario y, en consecuencia, pueden producir tallos de grosor sustancial (p. ej., palmas, árbol de Josué).

