
1.12: Sexo fúngico y grupos fúngicos
- Page ID
- 56504

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)
La estructura 'vegetativa' (es decir, no asociada con el sexo y la reproducción) de la mayoría de los hongos son generalmente muy consistentes: están compuestos por filamentos ramificados que exploran el volumen de material del que se alimentan (ya sea vivo o muerto) y esta estructura proporciona la adquisición de materiales que necesitan para crecer y sobrevivir. La consistencia de la forma vegetativa dificulta la clasificación de los hongos porque en su mayoría se parecen, aunque algunos varían en función de si los filamentos en coenocíticos o septados (con paredes transversales y por lo tanto celulares). Sin embargo, los hongos sí muestran variación, y pueden agruparse, en base a estructuras que están asociadas con la reproducción sexual. Las excepciones a esto son los dos grupos donde el sexo está completamente ausente (Glomeromycota—los hongos formadores de endomicorhizales) o raro (Quitridomicota—los quítridos). Los tres grupos restantes (mohos de pan = Zygomycota, hongos de copa = Ascomycota y hongos club = Basidiomycota generalmente presentan características específicas asociadas con la reproducción sexual que se utilizan definen estos grupos (generalmente considerados filos). Los hongos que no muestran rasgos sexuales solían ser puestos en un grupo llamado 'Fungi Imperfecti' (Deuteromycota), pero las técnicas moleculares ahora permiten que estos hongos se coloquen en uno de los grupos mencionados anteriormente.
Los hongos tienen las siguientes características asociadas con el sexo y la reproducción que son diferentes de los organismos sexuales más familiares:
- La producción a menudo involucra tanto aspectos sexuales como aspectos asexuales. Las células llamadas esporas, definidas como células con capacidades de dispersión (es decir, células móviles) y capacidades reproductivas (es decir, células que pueden convertirse en nuevos organismos fúngicos) a menudo son importantes tanto en la reproducción asexual como sexual.
- Los ametos G (células que pueden fusionarse con otros gametos) son bastante diferentes de lo que la mayoría consideraría 'típico'; suelen ser células hifas (parte de los filamentos fúngicos) que tienen la capacidad de fusionarse con otras hifas. Dependiendo del grupo, estas hifas especiales pueden tener o no características estructurales que las distinguirían de las hifas normales.
- La singamia a menudo implica dos pasos separados en el tiempo y a menudo en el espacio. Esto es consecuencia de que la fusión de las hifas que sirven como gametos generalmente no es seguida inmediatamente por la fusión de núcleos. La fusión celular se llama plasmogamia y la fusión nuclear se llama cariogamia. La plasmogamia seguida de una cariogamia retardada permite que los hongos tengan una nueva condición, el estado dikaryon, donde una célula tiene dos núcleos ('dikaryon' significa 'dos núcleos'), uno de cada progenitor. Esta condición a menudo se perpetúa: la célula del dicarión se divide mientras que ambos núcleos se dividen, formando así una nueva célula que también es dicarión. Este proceso puede continuar, produciendo múltiples células de dicarión e hifas de dikaryon. Obsérvese que aunque una célula dicariónica tiene dos copias de cada cromosoma, no se considera diploide porque cada núcleo es haploide, con solo una copia de cada cromosoma.
- En algún momento, algunas de las células diploides se vuelven diploides como resultado de la fusión de los dos núcleos (cariogamia) .Las células diploides NO se perpetúan; se someten a meiosis para formar núcleos haploides que desarrollan paredes para convertirse en células haploides. Así, el estado diploide es muy breve en extensión (es decir, tamaño y número de células) y a menudo breve en el tiempo también: las células diploides nunca se dividen mitóticamente para formar más células diploides, solo se dividen meióticamente para formar células haploides. Estas células haploides son, o pronto se convierten, esporas y se dispersan desde el hongo progenitor a una nueva ubicación donde germinan y forman hifas haploides.
- Las “cepas de apareamiento” son un mecanismo común que asegura que los hongos no se aparean consigo mismos. Dentro de una especie particular hay de dos a muchos tipos de apareamiento. Un tipo de apareamiento particular necesita encontrar un tipo de apareamiento diferente para interactuar sexualmente (es decir, fusionar hifas). La situación más simple tiene dos tipos de apareamiento: “+” y “-” y necesitan encontrarse para aparearse, pero puede haber más que solo dos tipos, por ejemplo, los tipos A, B, C, D, E y A podrían aparearse con cualquiera de los otros pero no con otro A. Generalmente, la interacción, o la falta de ella, implica señales químicas (feromonas) que son emitidos por un tipo de apareamiento y detectados únicamente por hongos de otro tipo de apareamiento. En respuesta a la feromona, las hifas crecen y se encuentran, provocando plasmogamia.
TEMAS
-
- Zygomycetes (moldes de pan)
- Ascomicetos (hongos de copa)
- Basidiomicetos (hongos club)
Zygomycetes, los moldes de pan


En los zigomicetos (Figura 2-3), la mayor reproducción es asexual y resulta de la producción de una estructura acechada (esporangióforo) terminada con esporangio esférico. Dentro del esporangio se encuentra el único tejido celular producido por el grupo, lo que significa que es una estructura donde los núcleos individuales se empaquetan uno a una célula. Estas células maduran en esporas que se dispersan cuando el esporangio se desintegra. Mucho menos frecuente es la reproducción asociada a un proceso sexual y a una estructura específica, la cigospora, de la que el grupo obtiene su nombre. El proceso se inicia cuando dos hifas compatibles perciben la presencia de la otra debido a las feromonas. Esto provoca que las hifas crezcan una hacia la otra y la producción de ramas hifas especializadas que son capaces de fusionarse entre sí. Dado que los moldes de pan son coenocíticos, la fusión de dos hifas (plasmogamia) permite que múltiples núcleos se acerquen (Fig 4). Inusual para el grupo, dos paredes transversales, una en cada una de las hifas fusionadas, se forman cerca del punto de fusión, creando una sola célula coenocítica con núcleos derivados de cada una de las dos cepas de apareamiento (Figura 5-6). Esta es la única célula de dicarión producida por los moldes de pan y se desarrolla en una estructura llamada zigospora (Figura 7-8) al agrandarse ligeramente y desarrollar una pared gruesa. Dentro de los núcleos haploides de zigospora de un tipo se emparejan connúcleos del otro tipo y se fusionan (cariogamia) para formar núcleos diploides, transformando la célula de ser dicarión a diploide, pero sigue siendo multinucleada. La cigospora generalmente está latente y generalmente tiene requisitos de germinación más específicos y una vida útil más larga que las esporas asexuales. Cuando la cigospora germina produce un esporangióforo comparable a los producidos asexualmente con un esporangio en su punta. A medida que esto se desarrolla, los núcleos diploides experimentan meiosis y los núcleos haploides se empaquetan individualmente en esporas a medida que se producen las paredes celulares. Las esporas se dispersan posteriormente, y cuando germinan producen hifas haploides coenocíticas. Obsérvese que el estado dikaryon está limitado a una sola célula y tiene una existencia muy breve.

Ascomycota — los hongos de copa

A diferencia de los mohos del pan, los hongos de copa (Ascomycetes), son septados, es decir, tienen paredes transversales y las 'hifas alimentadoras', las que adquieren nutrientes, están construidas de células que tienen un solo núcleo haploide. Muchos miembros del grupo, la reproducción es principalmente a exclusivamente asexual consecuencia de la producción de ramas hifales especializadas que producen pequeñas células (conidios) que se rompen fácilmente para servir como propágulos. Generalmente, estas células están latentes y tienen otras características celulares, por ejemplo, los alimentos almacenados, que promueven su papel en la reproducción y dispersión. Cuando y si se produce la reproducción sexual, se inicia cuando las cepas de apareamiento compatibles están cerca unas de otras y se comunican por feromonas. Se producen ramas hifales infladas, una en cada cepa, produciendo células que son ligeramente diferentes en forma y son multinucleadas. Cuando están cerca una de la otra, una de las células produce una extensión que toca la célula expandida del otro tipo de apareamiento y se fusiona con ella (se produce plasmogamia) y los núcleos haploides se mueven de una de las estructuras, denominada anteridio y considerada macho, a la otra, llamada ascogoniumand considerado femenino. Como resultado de la migración de núcleos, la célula de archegonio se convierte en dikaryon, con dos tipos de núcleos, uno de cada progenitor. El ascogonio desarrolla extensiones y dos núcleos (uno de cada tipo de apareamiento) se asocian y migran hacia ellos. Se forma una pared transversal, produciendo la primera celda de lo que se describe como hifa ascógena. Estas hifas crecen desde la punta y permanecen en dikaryon, con dos núcleos haploides por célula, resultado de una coordinación de expansión de la punta, dos divisiones nucleares (una por cada núcleo), migración del núcleo y formación de septos. El crecimiento de las hifas ascogneas más el crecimiento de ambas hifas parentales (haploides) producen un cuerpo fructífero de hifas densamente entrelazadas. El tamaño del cuerpo fructífero varía enormemente en tamaño, de aproximadamente 100 um a 10 cm o más, con la mayoría en el extremo más pequeño. El tamaño y la forma varían y pueden ser utilizados para identificar especies. El nombre común para el grupo (“hongo de copa”) se refiere a un cuerpo fructífero en forma de copa. Los hongos de copa se encuentran viviendo independientemente y también como el compañero fúngico de varios líquenes; muchos líquenes producen ascocarpos (Figura 10) que son relativamente grandes y visibles a simple vista. Otros cuerpos fructíferos tienen forma de matraces o completamente cerrados y típicamente tienen menos de un mm de extensión.

En una parte específica de este cuerpo fructífero se completa el ciclo sexual. Las puntas de las hifas ascógenas forman un gancho, volviéndose sobre sí misma y los dos núcleos se fusionan (se produce la cariogamia) en la célula que ha formado el gancho (crozier), convirtiéndola (brevemente) en una célula diploide. Las únicas células diploides que se encuentran en la Ascomycota son estas células específicas, eventualmente llamadas asci (singular = ascus) situadas en los extremos de las hifas ascógenas (estas hifas no producen células adicionales después de formar el gancho). La célula diploide se alarga y experimenta meiosis para formar una célula alargada con cuatro núcleos haploides. Cada uno de estos luego pasa por mitosis para producir una célula alargada llamada ascus con ocho núcleos haploides seguidos. Cada núcleo adquiere una pared celular y se desarrolla en lo que se llama ascosporas. Asci con ocho ascosporas son diagnósticos para el Ascomycota.

La presión hidrostática dentro del ascus hace que las ascosporas se dispersen por la fuerza cuando la punta del ascus se rompe. Las ascosporas dispersas germinan para formar hifas haploides, las cuales forman micelios haploides que pueden reproducirse asexualmente a través de conidios u otras esporas. Si un micelio haploide entra en contacto con hifas de otro tipo de apareamiento, se puede desencadenar la reproducción sexual.


Basidiomicetos — hongos club
Los hongos club son septados como los hongos de copa (Ascomycota). De todos los grupos fúngicoslos hongos club tienen el estado dikaryon más extenso. Como es el caso de casi todos los hongos, el estado dikaryon se inicia cuando dos hifas haploides y monocarionales compatibles se encuentran como resultado de la atracción química. Se produce plasmogamia, formando una célula de dicarión. Esta célula se divide y crece extensamente, formando un micelio de alimentación que es dikaryon. Muchos de los basidiomicota producen 'conexiones de pinza' (Figura 17) que pueden ayudar a mantener el estado de dicarión a medida que las hifas se alargan. Los núcleos sufren mitosis y se forman paredes transversales. A medida que la célula terminal se alarga, se forma un arco entre la célula terminal y su célula esparental. Cuando los dos núcleos se dividen, el arco permite que uno de los dos núcleos se mueva a la célula parental a medida que se forman septos. Esto asegura que tanto la célula hija como la célula madre tengan uno de cada tipo de núcleo.
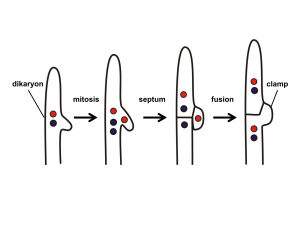
Recuérdeque solo hay una sola célula de dikaryon en los moldes de pan (Zygomycota) y las únicas células dikaryon de los hongos de copa (Ascomycota) se funden en los cuerpos fructíferos. Así, si uno encuentra una hifa dicariota fuera de un cuerpo fructífero entonces debe pertenecer a un hongo club (Basidiomycota). Las hifas dicariotas de los hongos club crecen y se alimentan extensamente hasta que se encuentran las condiciones adecuadas para desencadenar la 'fructificación'. En este punto, el patrón de crecimiento de al menos algunas de las hifas cambia de una donde se explora su sustrato para su nutrición a otra donde las hifas se entrelazan entre sí, formando la densa masa de hifas que se convertirán en cuerpo ruitante. A menudo, el cuerpo fructífero emerge del sustrato del que se alimenta el hongo, por ejemplo, un hongo que emerge del suelo o del tronco de un árbol. En algún lugar sobre o dentro de la densa masa de hifas se producen células especiales, denominadas basidios, que tienen “forma de palo” (Figura 18). Dentro de estas células se produce la cariogamia, transformándolas en células diploides. El núcleo diploide sufre meiosis para producir cuatro núcleos haploides, pero no se produce citocinesis, haciendo que el basidio sea 'cuadra-nucleado'. Cuatro extensiones crecen a partir del basidio y los cuatro núcleos migran a estos. A esto le sigue la formación de la pared celular para producir un basidio sin núcleo pero con cuatro células haploides poco unidas, llamadas basidiosporas, que se extienden desde ellas. Las esporas son liberadas balísticamente y son dispersadas por el viento. Cuando germinan forman nuevas hifas haploides.

A diferencia de los hongos de copa, cuyos cuerpos fructíferos son generalmente pequeños y no suelen notarse, los cuerpos fructíferos de muchos hongos club suelen ser relativamente grandes, 10-30 cm y tienen formas características que describimos como hongos (con tallo y capuchón) (Figura 19-20) u hongos de soporte ( aproximadamente en forma de pezuña y unida sin tallo a los troncos de las plantas leñosas). Dos patrones muy comunes de distribución de basidios son (1) extendiéndose desde finas aletas de tejido denominadas “branquias”, típicas del hongo comercialmente disponible, o (2) basidios que se extienden desde tubos (cilindros) de tejido que, cuando se ven en la superficie, aparecen como numerosos poros.





Dos grandes grupos de basidiomicetos, los tizones (Figura 22-23) (Clase Ustomicetos) y los oxiuros (Figura 24-25) (clase Teliomicetos) presentan patrones de sexo y reproducción ligeramente diferentes y a veces involucran patrones de sexo y reproducción asociados con su obligado estilo de vida parasitario. Muchos de estos organismos son muy importantes económicamente porque pueden reducir drásticamente los rendimientos de cultivos importantes. Tampoco forman 'cuerpos fructíferos', hechos únicamente de material fúngico, sino que provocan un crecimiento anormal (agallas) en la planta en la que están creciendo. Estas agallas están compuestas por células vegetales infectadas, células vegetales no infectadas que crecen anormalmente debido al parásito y algunas hifas fúngicas que interconectan las células infectadas. Si bien el basidiomiceto típico descrito anteriormente tiene un solo tipo de espora, las basidiosporas haploides formadas en el cuerpo fructífero, tanto las oxidas como las manchas suelen producir teliosporas, de una a varias esporas celulares, que contienen células dicariotas. Cuando las teliosporas germinan se produce la cariogamia, seguida de la meiosis, produciendo un basidio con núcleos haploides que producen basidiosporas haploidasporas.

Si bien la mayoría de los tizones solo producen estos dos tipos de esporas, las oxidaciones pueden tener dos o tres tipos más de esporas y un ciclo de vida complicado que a veces involucra a dos huéspedes. El ciclo de vida de la roya del trigo se muestra en la Figura 26 y se discute en el Capítulo 30.

Las basidiosporas infectan al huésped #1 (agracejo) y forman una estructura llamada picnia en la superficie superior de la hoja. La picnia produce dos estructuras que producen la singamia. Una de ellas son esporas haploides llamadas pincosporas (o espermatias) que podrían considerarse gametos. Las picnoesporas se liberan como células individuales en un dulce 'néctar' que atrae a los insectos que pueden transportarlas a otras picnias compatibles (es decir, de diferente tipo de apareamiento) donde pueden fusionarse (plasmogamia) con la segunda estructura producida por picnia involucrada en la singamia: 'hifas flexuosas' que se extienden fuera de la picnia. La fusión de la pincosporas con las células haploides de estas hifas provoca la singamia y forma la célula inicial de una hifa dikaryon que crece hasta el fondo de la hoja, forma una bola y libera esporas de dicarión (aeciosporas) que infectan al huésped #2 (trigo), provocando que forme otro tipo de esporas de dicarión, uridinosporas, que pueden infectar a más plantas de trigo. Al final de la temporada el tipo final de espora, la teliospora, se forma en el trigo. La cariogamia y la meiosis ocurren en las teliosporas y de ellas emergen promicelios (básicamente basidios) y producen basidiosporas.
