
1.15: Sexo y reproducción en plantas semilleras
- Page ID
- 56494

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Las semillas son una innovación notable que han sido muy importantes para la evolución de la vida vegetal. La gran mayoría de las plantas que observamos y utilizamos poseen semillas y las semillas dominan la mayoría de los hábitats terrestres. El último capítulo describía, en términos generales, qué son las semillas y qué modificaciones en el ciclo de vida general de la planta de alternancia de generaciones tuvieron que ocurrir para que las semillas aparecieran. Este capítulo rellena algunos de los detalles de los cinco grupos de plantas semilleras:
- plantas con flores, con 250, 000 especies, con mucho el grupo de plantas más diverso y ubicuo
- coníferas: aunque con solo 1000 especies, se encuentran muy comúnmente y son de importancia ecológica y económica
- cícadas: un pequeño grupo de alrededor de 300 especies con distribución e importancia limitadas
- gnetófitos: un pequeño grupo de solo tres géneros y alrededor de 100 especies
- Ginkgo: una sola especie que sobrevive solo donde se cultiva
Además de estos cinco grupos existentes, hay varios grupos de plantas semilleras bien representadas en fósiles pero que ya no están presentes. Estas especies extintas a veces se agrupan como 'helechos semilla', pero no se piensa que tanto el agrupamiento como la designación como grupo sean precisos: no están estrechamente alineados con los helechos y probablemente representan una agrupación polifilética. La mayoría de los trabajadores creen que las semillas evolucionaron más de una vez y por lo tanto que no debería haber ninguna entidad filogenética correspondiente a ninguna de las plantas de semillas ni a las gimnospermas (es decir, plantas de semillas que carecen de flores) aunque estas categorías persisten. Consideraremos los detalles del ciclo de vida, en particular la forma del gametofito femenino y los mecanismos asociados a la polinización y fertilización, para los cinco grupos de plantas semilleras existentes.
TEMAS
- Desarrollo de semillas de coníferas
- Desarrollo de semillas en otras gimnospermas
- Desarrollo de semillas en angiospermas
- Flores
- Modificaciones florales
- Frutas
Coníferas

Los pinos son las coníferas más comúnmente vistas y el grupo que se describirá a continuación pero el patrón básico se mantiene para todo el grupo. Las plantas que se reconocen como pinos son plantas diploides, esporófitas. Todas las coníferas, incluidos los pinos, son heterosporosas y producen dos tipos de esporas, ambas en un mismo árbol. Los sitios de producción de esporas son los conos. Los conos que la mayoría de la gente reconoce como 'conos de pino' son piñas hembra. Estos no son solo el sitio de producción de megasporas sino también el sitio de germinación de megasporas para formar un gametofito, producción de huevos por ese gametófito, fertilización de óvulos y desarrollo de semillas. Todos estos eventos tienen lugar en un lugar descrito como óvulo. Estos procesos generalmente toman varios años y las estructuras que uno suele reconocer como piñas han estado viviendo y desarrollándose durante un tiempo de dos años o más, con muchos de los eventos significativos ocurriendo cuando el cono es mucho más pequeño y no se observa tan fácilmente (Figura 1). Un cono hembra consiste en un eje (tallo) con escalas subtendidas por brácteas, con las escalas consideradas derivadas de ramas modificadas. Sobre la superficie superior de la escala se encuentran los óvulos, las estructuras que se desarrollan en semillas.

Las esporas masculinas se producen en conos de pino macho menos familiares, pero ciertamente fáciles de observar, que crecen más rápidamente que los conos femeninos pero que están presentes en el árbol por un tiempo mucho más corto. Por lo general, se producen en el otoño y son visibles como un racimo de estructuras en la base del brote. Estos se expanden en la primavera/principios del verano (Figura 2) y se secan y se marchitan un mes después. A diferencia de los conos femeninos, los conos m ale son de estructura simple: una rama con hojas fuertemente empaquetadas que llevan esporófilas, cada una con un par de esporangios relativamente grandes en su superficie inferior.
La generación de ófitos gametos se desarrolla a partir de microsporas y megasporas producidas y retenidas en los conos macho y hembra. Los gametofitos son muy reducidos y, especialmente para el gametofito femenino, en gran parte invisibles debido a su pequeño tamaño y ubicación. Los gametofitos masculinos se producen en las microsporangias de los conos masculinos. Estos inicialmente contienen células que sufren meiosis para producir microsporas. Al principio, las esporas se encuentran en grupos de cuatro que reflejan su origen en las dos divisiones de la meiosis (una célula a dos células por cuatro células). Las esporas finalmente se separan y experimentan un periodo de desarrollo muy limitado, produciendo un organismo haploide con cuatro núcleos, generalmente en tres células (es decir, una célula tiene dos núcleos), y poseen dos sacos de aire tipo ala (Figura 3). Particularmente significativo para el desarrollo del grano de polen (también conocido como gametofitos masculinos), es el hecho de que su desarrollo es detenido. Esto, y el hecho de que el microsporangio se abra, permite que el polen sea dispersado por el viento.
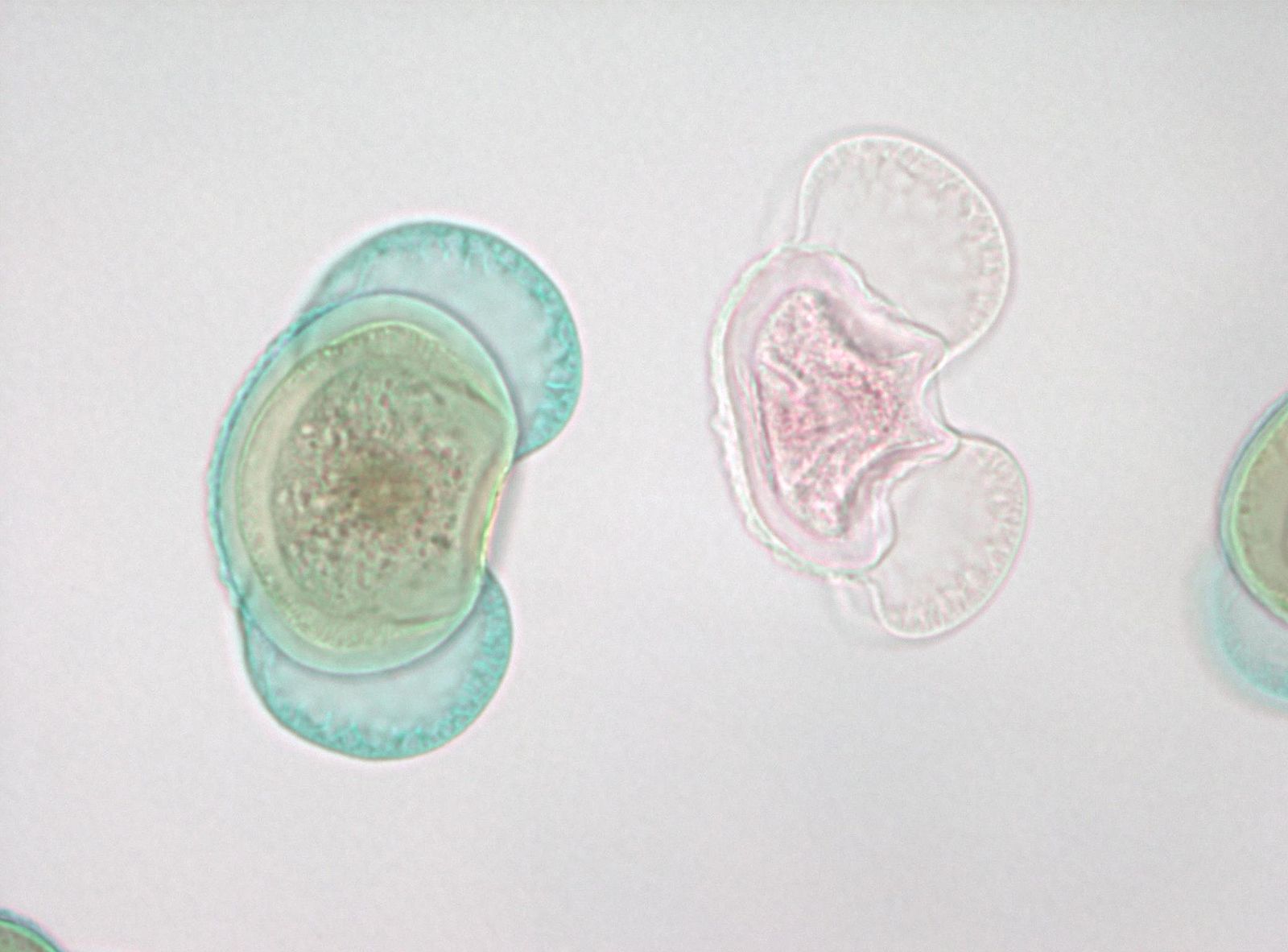
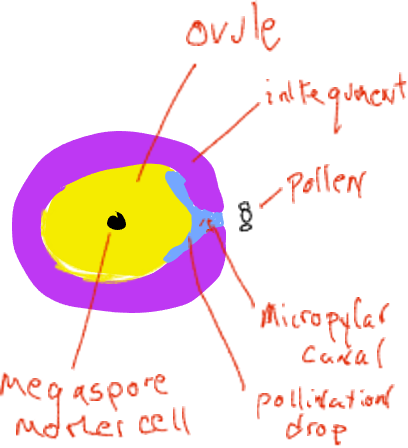
Polinización es el nombre de la transferencia del gametofito macho (polen) desde donde se produce a la ubicación del gametofito femenino, en coníferas, un movimiento de un cono de pino macho a una ubicación dentro del cono hembra. Al momento de la liberación de polen, los conos hembra son muy pequeños y están 'abiertos' con espacios por encima de cada escala de cono individual que están abiertos al exterior (Figura 4). Cada escama de cono hembra lleva en su superficie superior dos óvulos, cada uno con un megasporangio embebido en tejido esporofítico llamado tegumentos. Al principio del desarrollo del óvulo hay una abertura, el canal micropilar, entre los tegumentos que se conecta a un espacio entre las escamas del cono. Los granos de polen (gametofitos masculinos) en el aire pueden deslizarse entre las escamas del cono hembra y depositarse en el espacio próximo al canal micropilar. El óvulo secreta una 'gota de polinización' líquida en este espacio y los granos de polen terminan en el líquido y se rehidratan. En un mecanismo no completamente comprendido, el líquido con los granos de polen es retirado a través del canal micropilar a un espacio en el interior de las tegumentos adyacentes al megasporangio.

Poco después los tegumentos crecen para bloquear el canal micropilar y las escamas del cono crecen para sellar el cono hembra del exterior. En este momento el megasporangio tiene una sola célula madre megasporosa destinada a someterse a meiosis. Después de la meiosis, solo una de las cuatro células hijas permanece como megasporo. La megaspora no se dispersa sino que se desarrolla dentro del megasporangio en un gametofito femenino de varios miles de células que generalmente produce dos o tres archegonias, cada una de las cuales produce un solo huevo. Todo esto ocurre dentro del cono hembra que está unido y es parte de la planta esporofita. Los gametofitos femeninos de coníferas son 'organismos' altamente reducidos que se encuentran dentro de los conos ovulados, incrustados dentro del tejido esporofítico de la planta.
En el momento de la polinización no suele haber gametofitos femeninos, solo una megaspora. Dentro del cono, los gametofitos machos continúansu desarrollo, aunque muy lentamente en los pinos, requiriendo 12 meses entre polinización y fertilización.
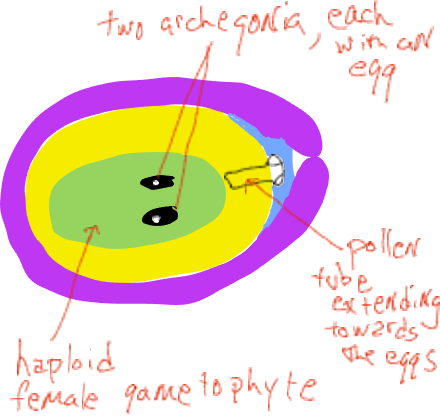
A medida que la megaspora produce una espora que lentamente se desarrolla en un gametofito femenino, pasa por una etapa de 'núcleo libre' donde las divisiones mitóticas no van acompañadas de formación de la pared celular (el organismo es coenocítico). Eventualmente se forman paredes celulares y el gametofito femenino forma estructuras descritas como archegonia, cada una con un solo óvulo.
A medida que se desarrolla el gametofito femenino, el polen germina y una sola célula se alarga desde el grano, creciendo a través del megasporangio (el nucello) hacia el gametofito femenino (Figura 6). Poco más de un año después de la polinización t su célula tubular se fusiona con el óvulo y se liberan dos núcleos espermáticos, uno de los cuales se fusiona con el núcleo del óvulo, formando un cigoto mientras que el otro núcleo se desintegra. Durante el tiempo entre la polinización y la fertilización, los conos femeninos crecen solo una pequeña cantidad y permanecen cerrados al exterior. Tenga en cuenta que no se produce esperma nadador, el gametofito masculino crece hasta el óvulo por medio de una célula alargada.
Después de la fertilización, el cigoto se convierte en embrión, embebido y nutrido por el gametofito femenino. Los tejidos que rodean al gametofito femenino se convierten en una capa de semilla, a menudo produciendo una estructura de ala que permite que la semilla sea dispersada por el viento. A medida que la semilla se desarrolla, el cono que la rodea también se desarrolla, a menudo creciendo sustancialmente. Después de la fertilización, las semillas pueden madurar en tan poco tiempo como un año, pero para la mayoría de las especies es de dos años o más. En la mayoría de los pinos, los conos finalmente se vuelven a abrir, permitiendo que las semillas se caigan y sean dispersadas por el viento. En ocasiones los conos permanecían cerrados y sólo abiertos tras el intenso calor de un fuego. Los conos hembra de enebros y tejos desarrollan características similares a frutas que atraen a los animales que facilitan la dispersión de semillas consumiendo los 'frutos' y defecando la semilla en una nueva ubicación.
Otras gimnospermas
Los otros tres grupos de plantas semilleras sin flores, Gnetófitos, Cícadas y Ginkgo presentan el mismo patrón básico de producción de semillas: las esporas masculinas se desarrollan en granos de polen que se dispersan del esporófito para terminar su desarrollo en la estructura que produce esporas femeninas y por lo tanto la hembra gametofitos. La polinización en al menos algunas cícadas y en algunos gnetófitos involucra insectos; en el ginkgo y la mayoría de los gnetófitos la polinización es por el viento. Gimnosperma significa literalmente 'semilla desnuda' y una característica que unifica a las plantas semilleras que no florecen es que, en el momento de la polinización, los óvulos son accesibles, no enterrados en tejidos por los que debe crecer el gametofito masculino; en cambio, los óvulosestán disponibles, al menos por un breve periodo de tiempo, debido a que las escamas de cono no se han fusionado entre sí y el canal micropilar está abierto. Sin embargo, el gametofito macho generalmente tiene que crecer a través del megasporangio (el nucello) para llegar al huevo. En las cícadas, el gametofito masculino en realidad desarrolla un tipo de estructura de alimentación (llamada haustorio), una estructura filamentosa ramificada que permea el nucelus y aparentemente obtiene alimento de él. Eventualmente se libera un espermatozoide flagelado y móvil que nada a través del líquido de una 'cámara archegonial', una zona de líquido entre el nucelo y el gametofito femenino. Los espermatozoides flagelados también se encuentran en el ginkgo.
Plantas con flores
El proceso básico de desarrollo de semillas en plantas con flores es el mismo que en las coníferas. Las principales diferencias incluyen las siguientes:
- Los órganos masculinos (productores de polen) y hembras (productores de semillas) generalmente se encuentran juntos en la misma estructura, la flor, no separados en dos ramas distintas como lo están en las coníferas.
- Los óvulos se producen dentro de una estructura llamada ovario, que no está abierta al exterior, requiriendo así que el gametofito masculino crezca a través de una distancia sustancial de tejido esporofítico para entrar en contacto con el gametofito femenino
- La transferencia de polen (polinización) a menudo involucra agentes biológicos (insectos, aves, raramente mamíferos) y una variedad de características florales mejoran la polinización.
- El gametofito femenino, que se llama saco embrionario (Figura 7).

Figura 7 Óvulo de angiosperma que contiene un gametofito femenino maduro (también conocido como saco embrionario). El gametofito contiene sólo siete células, una un óvulo que se fertiliza para formar un cigoto y una célula central grande que se fertiliza para formar el tejido del endospermo triploide. Las tegumentos circundantes se desarrollan en la capa de la semilla. debido a que finalmente contiene el embrión, es aún más educado que en otras plantas semilleras, drásticamente así, generalmente consiste en sólo siete células, seis células haploides, una de las cuales es el huevo, y una célula central más grande con dos núcleos haploides.
- Ambos espermnucleicosproducidos por el
gametofito masculino participan en un evento de fecundación (singamia). Uno se fusiona con el huevo para formar un cigoto y el segundo se fusiona con la célula central. Este núcleo espermático se combina con los dos núcleos de la célula central para formar un núcleo triploide 'endospermo'. La célula central entonces prolifera, formando un tejido, el endospermo, que tiene un desarrollo limitado y solo se encuentra durante el desarrollo de la semilla y a menudo, pero no siempre, en la semilla madura. Las etapas iniciales del desarrollo del endospermo involucran una etapa 'nuclear libre' donde los núcleos se dividen sin formación de pared celular, creando una célula multinucleada, toda triploide (coenocítica). Este material se llama 'endospermo líquido' y es familiar como la leche de coco, que en realidad es citosol. - En las angiospermas el gametofito femenino, que es muy limitado tanto en tamaño como en vida útil, no es el tejido nutritivo para el embrión en desarrollo como lo es en otras plantas semilleras, por ejemplo, las coníferas. En las angiospermas el tejido nutritivo para el embrión en desarrollo es el endospermo, el tejido resultante de un segundo evento de fertilización.
La flor

La flor es un tallo altamente modificado, típicamente con cuatro verticilos que se agrupan juntos al final de la rama. Se cree que los componentes de cada uno de los cuatro verticilos representan hojas modificadas, siendo los dos verticilos internos hojas portadoras de esporas altamente modificadas (esporófilas) (Figura 8-9). Los elementos del verticilo más externo (sépalos) son los más parecidos a las hojas, aunque a menudo bastante pequeños. Los elementos del siguiente verticilo (pétalos) suelen tener forma de hoja pero suelen ser estructuras grandes y coloridas que carecen de clorofila. El siguiente verticilo son s tamenos que a menudo consisten en un tallo (un filamento) que termina en una estructura llamada antera en la que se produce polen. Inicialmente, las anteras poseen microsporangios que contienen células madre de microsporas. Estas producen microsporas por meiosis y estas esporas germinan y se desarrollan en gametofitos masculinos, también conocidos como granos de polen, compuestos por solo dos o tres células. Cuando el polen está maduro la antera generalmente se abre para hacer que el polen sea accesible a los polinizadores o al viento. Los istilos P son el verticilo más interno y a menudo se fusionan entre sí, por lo que la estructura central de la flor es un pistilo único (compuesto). El pistilo (s) generalmente consiste en una base agrandada (el ovario), con una estructura acechada (el estilo) que emerge de su parte superior que termina con una superficie (el estigma) que recibe polen. Dentro del ovario se producen demasiados óvulos que eventualmente se convierten en semillas. Previo a esto, los óvulos son sitios de producción de megasporas, desarrollo de gametófitos femeninos (= saco embrionario), fertilización y finalmente el proceso de desarrollo de semillas. Después de la polinización, el gametofito macho de dos o tres celdas (polen) germina sobre el estigma y crece a través del estilo y luego obtiene acceso a un óvulo al crecer a través del canal micropilar, una abertura entre los tegumentos que rodean cada óvulo. El crecimiento del tubo de polen implica la expansión de una sola célula, llamada tubo de polen, que entrega dos gametos masculinos al extremo del saco embrionario con el óvulo . La célula tubular se fusiona con el saco embrionario y entrega dos espermatozoides para lograr una doble fertilización, permitiendo el desarrollo posterior del cigoto y el endospermo.

Modificaciones florales
Las variaciones en el plan básico de las flores son una de las historias asombrosas de la botánica y de toda la biología. En general, los cambios pueden atribuirse a las fuerzas de selección natural que actúan sobre la interacción entre polinizadores y plantas. Algunas de las transformaciones comunes del patrón descrito anteriormente incluyen:
- fusión de las partes de un verticilo, por ejemplo, todos los pétalos fusionados para formar una taza o embudo
- fusión de los miembros de dos verticilos (por ejemplo, fusión de estambres sobre los pétalos)
- reducción en el número de miembros de un verticilo, en particular una reducción de muchos pistilos a un solo pistilo como se encuentra en las flores de las flores de Asteraceae
- cambiar de simetría radial (siendo todas las partes de un verticilo del mismo tamaño y orientadas de manera similar, Figura 9-10) a simetría bilateral con flores que tienen dos lados que son imágenes especulares entre sí (Figura 11), o a veces para no tener simetría alguna.
- colocación del ovario por debajo del punto de unión de los otros verticilos
- eliminación de múltiples partes, formando a veces flores unisexuales (Figura 12).



El fruto
Después de la fertilización, el óvulo se transforma en un seedino en un proceso que implica el desarrollo coordinado de tres tejidos distintos:
- el cigoto se convierte en un embrión;
- el endospermo prolifera, primero en un patrón “nuclear libre” (las divisiones nucleares no van acompañadas de formación de la pared celular) y posteriormente produciendo nuevas células, y finalmente, en algunos grupos, el endospermo desaparece a medida que el embrión agranda su (s) cotiledón (s);
- los tejidos que rodean el embrión y el endospermo se convierten en una capa de semilla.
Obsérvese nuevamente que la composición genética de estos tres componentes difiere: el embrión es una 'nueva generación' y es diploide, el endospermo es triploide y los tejidos circundantes son diploides pero son de una generación anterior a la del embrión.
Mientras que el óvulo se transforma en una semilla, el ovario, y a veces otros tejidos que rodean el ovario, se desarrollan en una estructura llamada fruto. La transformación del ovario en un fruto generalmente implica la producción de nuevas células, el crecimiento de estas células y el desarrollo de características específicas de la planta que se observan. El fruto generalmente tiene características que mejoran la dispersión de las semillas y a menudo tiene características que protegen la semilla. Aunque generalmente es fácil distinguir la semilla del fruto, ocasionalmente no hay una demarcación obvia entre ellas, puede haber una demarcación que sea engañosa. Por ejemplo, las almendras se derivan de una fruta que es como una cereza (de hecho las cerezas y las almendras están muy estrechamente relacionadas). Comemos la parte carnosa de las cerezas, que forma parte del fruto. El 'hoyo' no es en realidad una semilla sino más bien es la semilla rodeada por la capa más interna del fruto. Cuando 'cáscara' una almendra, está rompiendo el tejido de la fruta abierta para revelar una sola semilla en su interior. En este caso el papel protector de la cubierta de la semilla ha sido asumido por una porción del fruto (Figura 13). De manera similar, la 'semilla' de asunflor es en realidad un fruto de una sola semilla y el bombardeo de la 'semilla' en realidad está dividiendo el fruto (Figura 14).